Kieselalgen
| Kieselalgen | |
|---|---|

| |
| Lichtmikroskopie einer Probe mariner Kieselalgen, die zwischen den Kristallen des jährlichen Meereises in der Antarktis leben und eine Vielzahl von Größen und Formen aufweisen | |
| Wissenschaftliche Klassifizierung | |
| Klade: | SAR |
| Infrakönigreich: | Heterokonta |
| Phylum: | Ochrophyta |
| Unterstamm: | Khakista |
| Klasse: | Bacillariophyceae Dangeard, 1933 |
| Synonyme | |
| |
Kieselalgen (neulateinisch diatoma) sind Mitglieder einer großen Gruppe, die mehrere Gattungen von Algen, insbesondere Mikroalgen, umfasst und in den Ozeanen, Gewässern und Böden der Welt vorkommt. Lebende Kieselalgen machen einen beträchtlichen Teil der Biomasse der Erde aus: Sie erzeugen etwa 20 bis 50 Prozent des jährlich auf dem Planeten produzierten Sauerstoffs, nehmen jedes Jahr über 6,7 Milliarden Tonnen Silizium aus den Gewässern auf, in denen sie leben, und machen fast die Hälfte des organischen Materials in den Ozeanen aus. Die Schalen toter Kieselalgen können bis zu 800 m tief auf dem Meeresboden liegen, und das gesamte Amazonasbecken wird jährlich mit 27 Millionen Tonnen Kieselalgenschalenstaub gedüngt, der von transatlantischen Winden aus der afrikanischen Sahara herantransportiert wird, ein Großteil davon aus der Bodélé-Senke, die einst aus einem System von Süßwasserseen bestand. ⓘ
Kieselalgen sind einzellige Organismen: Sie kommen entweder als Einzelzellen oder in Kolonien vor, die die Form von Bändern, Fächern, Zickzacklinien oder Sternen annehmen können. Die Größe der einzelnen Zellen liegt zwischen 2 und 200 Mikrometern. Bei Vorhandensein ausreichender Nährstoffe und Sonnenlicht verdoppelt sich eine Ansammlung lebender Kieselalgen etwa alle 24 Stunden durch ungeschlechtliche Mehrfachspaltung; die maximale Lebensdauer einzelner Zellen beträgt etwa sechs Tage. Kieselalgen haben zwei unterschiedliche Formen: einige wenige (zentrische Kieselalgen) sind radialsymmetrisch, während die meisten (pennate Kieselalgen) weitgehend bilateral symmetrisch sind. Ein einzigartiges Merkmal der Anatomie der Kieselalgen ist, dass sie von einer Zellwand aus Siliziumdioxid (hydratisiertes Siliziumdioxid) umgeben sind, die als Kegelstumpf bezeichnet wird. Aufgrund ihrer photonischen Nanostruktur besitzen diese Frusteln eine strukturelle Färbung, weshalb sie auch als "Juwelen des Meeres" und "lebende Opale" bezeichnet werden. Die Bewegung der Kieselalgen erfolgt in erster Linie passiv durch Meeresströmungen und windbedingte Wasserturbulenzen; die männlichen Gameten der zentrischen Kieselalgen besitzen jedoch Geißeln, die eine aktive Bewegung bei der Suche nach den weiblichen Gameten ermöglichen. Ähnlich wie Pflanzen wandeln Diatomeen Lichtenergie durch Photosynthese in chemische Energie um, aber ihre Chloroplasten wurden auf unterschiedliche Weise erworben. ⓘ
Ungewöhnlich für autotrophe Organismen verfügen Diatomeen über einen Harnstoffzyklus, ein Merkmal, das sie mit den Tieren teilen, obwohl dieser Zyklus bei Diatomeen zu anderen Stoffwechselzwecken genutzt wird. Die Familie der Rhopalodiaceae besitzt auch einen cyanobakteriellen Endosymbionten, den sogenannten Sphäroidkörper. Dieser Endosymbiont hat seine photosynthetischen Eigenschaften verloren, aber seine Fähigkeit zur Stickstofffixierung beibehalten, so dass die Kieselalge atmosphärischen Stickstoff fixieren kann. Andere Kieselalgen, die mit stickstofffixierenden Cyanobakterien eine Symbiose eingehen, gehören zu den Gattungen Hemiaulus, Rhizosolenia und Chaetoceros. ⓘ
Das Studium der Kieselalgen ist ein Teilgebiet der Phykologie. Kieselalgen gehören zu den Eukaryonten, d. h. zu den Organismen mit einem Zellkern mit Kernhülle, was sie von den Prokaryonten Archaeen und Bakterien unterscheidet. Diatomeen gehören zu den Planktonarten, die als Phytoplankton bezeichnet werden und am häufigsten vorkommen. Diatomeen wachsen auch an benthischen Substraten, auf Treibgut und auf Makrophyten. Sie sind ein wesentlicher Bestandteil der Periphyton-Gemeinschaft. Eine andere Klassifizierung unterteilt das Plankton nach seiner Größe in acht Arten: In diesem Schema werden Kieselalgen zu den Mikroalgen gezählt. Es gibt mehrere Systeme zur Klassifizierung der einzelnen Kieselalgenarten. ⓘ
Fossile Beweise deuten darauf hin, dass Kieselalgen während oder vor der frühen Jurazeit, also vor etwa 150 bis 200 Millionen Jahren, entstanden sind. Der älteste fossile Nachweis für Kieselalgen ist ein Exemplar der heute lebenden Gattung Hemiaulus in spätjurassischem Bernstein aus Thailand. ⓘ
Kieselalgen werden zur Überwachung vergangener und gegenwärtiger Umweltbedingungen verwendet und werden häufig in Studien zur Wasserqualität eingesetzt. Kieselgur (Kieselgur) ist eine Sammlung von Kieselalgenschalen, die in der Erdkruste vorkommen. Es handelt sich um weiche, kieselsäurehaltige Sedimentgesteine, die sich leicht zu einem feinen Pulver zerbröseln lassen und in der Regel eine Partikelgröße von 10 bis 200 μm aufweisen. Kieselgur wird für eine Vielzahl von Zwecken verwendet, u. a. zur Wasserfilterung, als mildes Schleifmittel, in Katzenstreu und als Dynamitstabilisator. ⓘ
Zeigt Überlagerungen aus vier Fluoreszenzkanälen
(b) Cyan: [PLL-A546-Fluoreszenz] - generische Gegenfärbung zur Visualisierung eukaryotischer Zelloberflächen
(c) Blau: [Hoechst-Fluoreszenz] - färbt DNA an, identifiziert Zellkerne
(d) Rot: [Chlorophyll-Autofluoreszenz] - macht Chloroplasten sichtbar
| Kieselalgen ⓘ | ||||||||
|---|---|---|---|---|---|---|---|---|
|
Diese marinen Kieselalgen leben als „Eisalgen“ im Inneren des Meereises des antarktischen McMurdo-Sunds. | ||||||||
| Systematik | ||||||||
| ||||||||
| Wissenschaftlicher Name | ||||||||
| Bacillariophyta | ||||||||
| Haeckel |
.jpg)
Die Kieselalgen oder Diatomeen (Bacillariophyta) bilden ein Taxon von Photosynthese betreibenden Protisten (Protista) und werden in die Gruppe der Stramenopilen (Stramenopiles) eingeordnet. ⓘ
Oft wird die Gruppe mit dem synonymen Namen Diatomea Dumortier bezeichnet, alternativ sind auch die synonymen Namen Fragilariophyceae, Diatomophyceae in Verwendung. Einige Autoren nennen die Kieselalgen Bacillariophyceae, sie ordnen sie also als Klasse in die, dann als Phylum aufgefassten photosynthetischen Vertreter der Stramenopiles (Ochrophyta) ein, diese Auffassung wird etwa in den Datenbanken DiatomBase und WoRMS vertreten. Diese Verwendung ist allerdings missverständlich, da andere Taxonomen eine enger abgegrenzte Klasse Bacillariophyceae, als eine von drei Klassen innerhalb der Diatomeen aufführen. Bei Verwendung dieses Namens ist also die jeweilige Auffassung zu kontrollieren, da es sonst zu Missverständnissen kommt. ⓘ
Man unterscheidet heute rund 6000 Arten. Es wird jedoch angenommen, dass insgesamt bis zu 100.000 Arten existieren. Eine Arbeitsstelle für Diatomeen-Forschung (mit umfangreicher Sammlung und Online-Katalog), begründet von Friedrich Hustedt, befindet sich im Alfred-Wegener-Institut. ⓘ
Morphologie
Kieselalgen sind in der Regel 2 bis 200 Mikrometer groß, wobei es einige größere Arten gibt. Ihre gelblich-braunen Chloroplasten, der Ort der Photosynthese, sind typisch für Heterokonten, haben vier Zellmembranen und enthalten Pigmente wie das Carotinoid Fucoxanthin. Die Individuen haben in der Regel keine Geißeln, aber in den männlichen Gameten der zentrischen Diatomeen sind sie vorhanden und haben die übliche Heterokont-Struktur, einschließlich der für andere Gruppen charakteristischen Haare (Mastigoneme). ⓘ
Kieselalgen werden aufgrund ihrer optischen Eigenschaften oft als "Juwelen des Meeres" oder "lebende Opale" bezeichnet. Die biologische Funktion dieser strukturellen Färbung ist nicht klar, aber es wird vermutet, dass sie mit Kommunikation, Tarnung, Wärmeaustausch und/oder UV-Schutz zusammenhängen könnte. ⓘ
Kieselalgen bilden komplizierte harte, aber poröse Zellwände, die sogenannten Frusteln, die hauptsächlich aus Kieselsäure bestehen. Diese Kieselwand kann stark gemustert sein, mit einer Vielzahl von Poren, Rippen, winzigen Stacheln, Randleisten und Erhebungen, die alle zur Abgrenzung von Gattungen und Arten verwendet werden können. ⓘ
Die Zelle selbst besteht aus zwei Hälften, von denen jede eine im Wesentlichen flache Platte oder Klappe und ein marginales Verbindungs- oder Gürtelband enthält. Die eine Hälfte, die Hypothek, ist etwas kleiner als die andere Hälfte, die Epithek. Die Morphologie der Kieselalgen ist unterschiedlich. Obwohl die Form der Zelle typischerweise kreisförmig ist, können einige Zellen dreieckig, quadratisch oder elliptisch sein. Ihr Erkennungsmerkmal ist eine harte Mineralschale oder ein Kegelstumpf aus Opal (hydratisierte, polymerisierte Kieselsäure). ⓘ

2) Nukleolus; Sitz der Chromosomen
3) Golgi-Apparat; modifiziert Proteine und schickt sie aus der Zelle heraus
4) Zellwand; äußere Membran der Zelle
5) Pyrenoid; Zentrum der Kohlenstofffixierung
6) Chromatophor; pigmenttragende Membranstruktur
7) Vakuolen; Bläschen einer Zelle, die eine durch eine Membran gebundene Flüssigkeit enthalten
8) Zytoplasmatische Stränge; enthalten den Zellkern
9) Mitochondrien; erzeugen ATP (Energie) für die Zelle
10) Ventile/Striae; lassen Nährstoffe in die Zelle hinein und Abfallstoffe aus ihr heraus ⓘ

b) Striae (Poren, Pünktchen, Flecken oder Punkte in einer Linie auf der Oberfläche)
c) Raphe (Schlitz in den Klappen)
d) Zentralknötchen (Verdickung der Wand in der Mitte der Raphe)
e) Stigmata (Löcher durch die Klappenoberfläche, die außen rund, innen aber schlitzförmig aussehen)
f) Punctae (Flecken oder kleine Perforationen auf der Oberfläche)
g) Polarknötchen (Verdickung der Wand an den distalen Enden der Raphe) ⓘ

Kieselalgen werden in zwei Gruppen eingeteilt, die sich durch die Form der Kegelstümpfe unterscheiden: die zentrischen Kieselalgen und die pennaten Kieselalgen. ⓘ
Die pennaten Diatomeen sind bilateralsymmetrisch. Ihre Klappen haben Öffnungen in Form von Schlitzen entlang der Rippen, und ihre Schalen sind in der Regel parallel zu diesen Rippen langgestreckt. Sie erzeugen Zellbewegung durch Zytoplasma, das entlang der Raphien strömt und sich immer entlang fester Oberflächen bewegt. ⓘ
Zentrische Diatomeen sind radialsymmetrisch. Sie bestehen aus einer oberen und einer unteren Klappe - Epitheca und Hypotheca -, die jeweils aus einer Klappe und einem Gürtelband bestehen, die leicht untereinander gleiten und sich ausdehnen können, um den Zellinhalt im Laufe der Entwicklung der Diatomeen zu vergrößern. Das Zytoplasma der zentrischen Diatomee befindet sich entlang der inneren Oberfläche der Schale und bildet eine hohle Auskleidung um die große Vakuole in der Mitte der Zelle. Diese große, zentrale Vakuole ist mit einer Flüssigkeit gefüllt, die als "Zellsaft" bezeichnet wird und dem Meerwasser ähnelt, aber je nach spezifischem Ionengehalt unterschiedlich ist. Die Zytoplasmaschicht beherbergt mehrere Organellen, wie die Chloroplasten und Mitochondrien. Bevor die zentrische Diatomee zu expandieren beginnt, befindet sich ihr Zellkern in der Mitte einer der Klappen und beginnt, sich zur Mitte der Zytoplasmaschicht zu bewegen, bevor die Teilung abgeschlossen ist. Zentrische Kieselalgen haben eine Vielzahl von Formen und Größen, je nachdem, von welcher Achse aus sich die Schale erstreckt und ob Stacheln vorhanden sind. ⓘ


Verhalten
Die meisten zentrischen und araphiden pennaten Kieselalgen sind unbeweglich, und ihre relativ dichten Zellwände lassen sie leicht sinken. Planktonische Formen in offenen Gewässern sind in der Regel auf die turbulente Durchmischung der oberen Schichten des Ozeanwassers durch den Wind angewiesen, damit sie im sonnenbeschienenen Oberflächenwasser in der Schwebe bleiben. Viele planktische Diatomeen haben auch Merkmale entwickelt, die ihre Sinkgeschwindigkeit verlangsamen, wie z. B. Stacheln oder die Fähigkeit, in Kolonieketten zu wachsen. Diese Anpassungen erhöhen das Verhältnis von Oberfläche zu Volumen und den Widerstand, so dass sie länger in der Wassersäule schweben können. Einzelne Zellen können den Auftrieb über eine Ionenpumpe regulieren. ⓘ
Einige pennate Diatomeen sind zu einer Art der Fortbewegung fähig, die als "Gleiten" bezeichnet wird und die es ihnen ermöglicht, sich über Oberflächen mittels klebrigem Schleim zu bewegen, der durch die Raphe (ein länglicher Schlitz in der Ventilfläche) abgesondert wird. Damit eine Diatomeenzelle gleiten kann, muss sie ein festes Substrat haben, an dem der Schleim haften kann. ⓘ
Die Zellen stehen einzeln oder sind zu Kolonien verschiedener Art zusammengeschlossen, die durch kieselige Strukturen, Schleimpolster, -stängel oder -röhren, amorphe Schleimmassen oder durch Chitinfäden (Polysaccharid), die durch Streben der Zelle abgesondert werden, miteinander verbunden sein können. ⓘ
_(20671468900)-47.jpg)
Gran, 1897
_(20671468900)-48%2B49.jpg)
_(20671468900)-cropped.jpg)

Diese Projektion eines Stapels konfokaler Bilder zeigt die Zellwand der Diatomeen (cyan), Chloroplasten (rot), DNA (blau), Membranen und Organellen (grün). ⓘ
Lebenszyklus


Fortpflanzung und Zellgröße
Die Fortpflanzung bei diesen Organismen erfolgt ungeschlechtlich durch binäre Spaltung, bei der sich die Kieselalge in zwei Teile teilt und zwei "neue" Kieselalgen mit identischen Genen entstehen. Jeder neue Organismus erhält eine der beiden Keimblätter - ein größeres und ein kleineres - des Elternteils, das nun als Epithek bezeichnet wird, und wird zum Aufbau eines zweiten, kleineren Keimblattes, der Hypothek, verwendet. Die Diatomee, die den größeren Kegelstumpf erhalten hat, wird genauso groß wie ihr Elternteil, aber die Diatomee, die den kleineren Kegelstumpf erhalten hat, bleibt kleiner als ihr Elternteil. Dies führt dazu, dass die durchschnittliche Zellgröße dieser Kieselalgenpopulation abnimmt. Es wurde jedoch beobachtet, dass bestimmte Taxa die Fähigkeit haben, sich zu teilen, ohne eine Verringerung der Zellgröße zu verursachen. Um jedoch die Zellgröße einer Kieselalgenpopulation wiederherzustellen, müssen sexuelle Fortpflanzung und Auxosporenbildung stattfinden. ⓘ
Zellteilung

Die Diatomeen sind diploid und vermehren sich hauptsächlich ungeschlechtlich durch Zellteilung. Die Tochterzellen erhalten jeweils einen Schalenteil und bilden den anderen Teil neu; hiervon leitet sich auch die Bezeichnung „Diatomee“ (altgriechisch διατέμνειν (diatemnein) = spalten) ab. Der neue Schalenteil ist stets die kleinere Hypotheka, so dass im Generationenverlauf die Zellgröße fast aller Nachkommen fortlaufend schwindet, nur die Tochterzelllinie der Ausgangs-Epitheka behält die ursprüngliche maximale Größe bei. Wird eine Minimalgröße unterschritten, stirbt das Individuum. Bevor eine Minimalgröße erreicht wird, können jedoch Sexualvorgänge stattfinden. Aus den Zellen bilden sich durch Meiose haploide Gameten. Bei zentrischen Kieselalgen wurde Oogamie nachgewiesen: Die Gameten werden frei, nach Verschmelzen eines weiblichen mit einem männlichen Gameten bildet sich aus der Zygote unter Größenwachstum eine Dauerform, eine sogenannte Auxospore. Bei pennaten Kieselalgen wurde Konjugation beobachtet: Zwei Partner legen sich aneinander und bilden eine gemeinsame Cytoplasmabrücke („Konjugationskanal“), in die jeweils ein haploider Kern und ein Chloroplast der beiden Partner einwandern. Aus der so gebildeten Zygote bildet sich eine Auxospore, in der die Kernverschmelzung (Karyogamie) stattfindet. Aus den Auxosporen der zentrischen und pennaten Kieselalgen wird jeweils eine größere neue Kieselalge mit einer neuen zweiteiligen Schale gebildet. ⓘ
Beweglichkeit der Spermien
Diatomeen sind meist unbeweglich; bei einigen Arten können die Spermien jedoch geflaggt sein, wobei sich die Beweglichkeit in der Regel auf eine Gleitbewegung beschränkt. Bei zentrischen Diatomeen haben die kleinen männlichen Geschlechtszellen eine Geißel, während die weiblichen Geschlechtszellen groß und unbeweglich sind (oogam). Bei den pennaten Diatomeen hingegen haben beide Gameten keine Geißeln (isogam). Bestimmte araphide Arten, d. h. pennate Diatomeen ohne Raphe (Saum), sind als anisogam beschrieben worden und werden daher als Übergangsstadium zwischen zentrischen und raphiden pennaten Diatomeen, Diatomeen mit Raphe, angesehen. ⓘ
Zersetzung durch Mikroben
Bestimmte Bakterienarten in Meeren und Seen können die Auflösung von Kieselsäure in toten und lebenden Kieselalgen beschleunigen, indem sie hydrolytische Enzyme einsetzen, um das organische Algenmaterial abzubauen. ⓘ
Ökologie


Die Flüsse sind in T mol Si y-1 (28 Millionen Tonnen Silizium pro Jahr) angegeben. ⓘ


gegen Silikatkonzentration ⓘ
Verbreitung
Kieselalgen sind eine weit verbreitete Gruppe, die in den Ozeanen, im Süßwasser, in Böden und auf feuchten Oberflächen vorkommt. In nährstoffreichen Küstengewässern und während der ozeanischen Frühjahrsblüte sind sie eine der dominierenden Komponenten des Phytoplanktons, da sie sich schneller teilen können als andere Phytoplanktongruppen. Die meisten leben pelagisch im offenen Wasser, einige aber auch als Oberflächenfilme an der Wasser-Sediment-Grenzfläche (benthisch) oder sogar unter feuchten atmosphärischen Bedingungen. Sie sind besonders wichtig in den Ozeanen, wo sie schätzungsweise 45 % der gesamten ozeanischen Primärproduktion an organischem Material beitragen. Die räumliche Verteilung der marinen Phytoplanktonarten ist sowohl horizontal als auch vertikal begrenzt. ⓘ
Wachstum
Planktonische Diatomeen in Süßwasser- und Meeresumgebungen weisen typischerweise eine "Boom-and-Bust"-Lebensweise (oder "Bloom-and-Bust"-Lebensweise) auf. Wenn die Bedingungen in der oberen Mischschicht (Nährstoffe und Licht) günstig sind (wie im Frühjahr), können sie aufgrund ihres Wettbewerbsvorteils und ihrer schnellen Wachstumsrate die Phytoplanktongemeinschaften dominieren ("Boom" oder "Bloom"). Als solche werden sie oft als opportunistische r-Strategen eingestuft (d. h. als Organismen, deren Ökologie durch eine hohe Wachstumsrate r gekennzeichnet ist). ⓘ
Beitrag zum modernen ozeanischen Siliziumkreislauf
Kieselalgen tragen in erheblichem Maße zum modernen ozeanischen Siliziumkreislauf bei: Sie sind die Quelle für den größten Teil der biologischen Produktion. ⓘ
Auswirkungen
Die Süßwasserkieselalge Didymosphenia geminata, gemeinhin als Didymo bekannt, verursacht in Wasserläufen, in denen sie blüht, schwere Umweltschäden, da sie große Mengen eines braunen gallertartigen Materials produziert, das als "brauner Rotz" oder "Steinrot" bezeichnet wird. Diese Kieselalge ist in Europa beheimatet und gilt sowohl auf den Antipoden als auch in Teilen Nordamerikas als invasive Art. Das Problem wird am häufigsten aus Australien und Neuseeland gemeldet. ⓘ
Wenn die Bedingungen ungünstig werden, in der Regel bei Nährstoffmangel, sinken die Kieselalgenzellen in der Regel schneller ab und verlassen die obere Mischschicht ("bust"). Dieses Absinken wird entweder durch den Verlust der Auftriebskontrolle, die Synthese von Schleim, der die Kieselalgenzellen zusammenklebt, oder die Produktion von schweren, ruhenden Sporen ausgelöst. Durch das Absinken aus der oberen gemischten Schicht werden die Diatomeen den für ihr Wachstum ungünstigen Bedingungen entzogen, einschließlich Weidegängerpopulationen und höheren Temperaturen (die andernfalls den Zellstoffwechsel erhöhen würden). Zellen, die tieferes Wasser oder den flachen Meeresboden erreichen, können dann ruhen, bis die Bedingungen wieder günstiger werden. Im offenen Ozean gehen viele absinkende Zellen in die Tiefe verloren, aber in der Nähe der Sprungschicht können Rückzugspopulationen überleben. ⓘ
Schließlich gelangen die Kieselalgenzellen dieser Ruhepopulationen wieder in die obere gemischte Schicht, wenn sie von der vertikalen Durchmischung mitgerissen werden. In den meisten Fällen werden durch diese Durchmischung auch die Nährstoffe in der oberen gemischten Schicht wieder aufgefüllt, was den Boden für die nächste Runde der Kieselalgenblüte bereitet. Im offenen Ozean (abseits von Gebieten mit kontinuierlichem Auftrieb) findet dieser Zyklus aus Blüte, Büste und Rückkehr zu den Bedingungen vor der Blüte in der Regel in einem Jahreszyklus statt, wobei Kieselalgen nur im Frühjahr und Frühsommer vorherrschen. An einigen Orten kann es jedoch zu einer Herbstblüte kommen, die durch den Zusammenbruch der Sommerschichtung und den Eintrag von Nährstoffen verursacht wird, während die Lichtverhältnisse noch ausreichend für das Wachstum sind. Da die vertikale Durchmischung zunimmt und die Lichtverhältnisse mit dem Herannahen des Winters abnehmen, sind diese Blüten kleiner und von kürzerer Lebensdauer als ihre Pendants im Frühjahr. ⓘ
Im offenen Ozean wird die Kieselalgenblüte (im Frühjahr) in der Regel durch einen Mangel an Silizium beendet. Im Gegensatz zu anderen Mineralien ist der Bedarf an Silizium nur bei Kieselalgen gegeben, und es wird im Plankton-Ökosystem nicht so effizient regeneriert wie beispielsweise Stickstoff- oder Phosphornährstoffe. Dies wird in Karten der Nährstoffkonzentrationen an der Oberfläche deutlich - wenn die Nährstoffe entlang von Gradienten abnehmen, ist Silizium in der Regel als erstes erschöpft (normalerweise gefolgt von Stickstoff und Phosphor). ⓘ
Aufgrund dieses Zyklus von Blüte und Erschöpfung wird angenommen, dass Kieselalgen eine unverhältnismäßig wichtige Rolle bei der Ausfuhr von Kohlenstoff aus den Oberflächengewässern der Ozeane spielen (siehe auch die biologische Pumpe). Bezeichnenderweise spielen sie auch eine Schlüsselrolle bei der Regulierung des biogeochemischen Zyklus von Silizium im modernen Ozean. ⓘ
Grund für den Erfolg
Kieselalgen sind ökologisch erfolgreich und kommen in praktisch jeder Umgebung vor, die Wasser enthält - nicht nur in Ozeanen, Meeren, Seen und Flüssen, sondern auch in Böden und Feuchtgebieten. Viele Forscher sind der Meinung, dass die Verwendung von Silizium durch Kieselalgen der Schlüssel zu diesem ökologischen Erfolg ist. Raven (1983) wies darauf hin, dass die Synthese von Siliciumdioxidfruchtkörpern im Vergleich zu organischen Zellwänden weniger Energie erfordert (etwa 8 % einer vergleichbaren organischen Wand), was eine erhebliche Einsparung im Gesamtenergiehaushalt der Zelle bedeuten könnte. In einer mittlerweile klassischen Studie stellten Egge und Aksnes (1992) fest, dass die Dominanz von Kieselalgen in Mesokosmengemeinschaften in direktem Zusammenhang mit der Verfügbarkeit von Kieselsäure steht - bei Konzentrationen von mehr als 2 μmol m-3 stellten sie fest, dass Kieselalgen in der Regel mehr als 70 % der Phytoplanktongemeinschaft ausmachen. Andere Forscher vermuten, dass die biogene Kieselsäure in den Zellwänden der Diatomeen als wirksamer pH-Puffer wirkt und die Umwandlung von Bikarbonat in gelöstes CO2 (das leichter assimiliert wird) erleichtert. Ungeachtet dieser möglichen Vorteile, die sich aus der Verwendung von Silizium ergeben, haben Kieselalgen in der Regel höhere Wachstumsraten als andere Algen der gleichen Größe. ⓘ
Quellen für die Sammlung
Kieselalgen können aus verschiedenen Quellen gewonnen werden. Marine Kieselalgen können durch direkte Wasserprobenahme gesammelt werden, und benthische Formen können durch Abschaben von Seepocken, Austern- und anderen Schalen gewonnen werden. Kieselalgen sind häufig als brauner, glitschiger Belag auf untergetauchten Steinen und Stöcken zu finden und können mit der Flussströmung "fließen". Im Oberflächenschlamm eines Teichs, Grabens oder einer Lagune sind fast immer einige Kieselalgen zu finden. Lebende Kieselalgen finden sich oft in großer Zahl an fadenförmigen Algen oder bilden gallertartige Massen auf verschiedenen untergetauchten Pflanzen. Cladophora ist häufig mit Cocconeis, einer elliptisch geformten Kieselalge, bedeckt; Vaucheria ist oft mit kleinen Formen bedeckt. Da Kieselalgen ein wichtiger Bestandteil der Nahrung von Weichtieren, Manteltieren und Fischen sind, finden sich im Verdauungstrakt dieser Tiere oft Formen, die auf andere Weise nicht leicht zu sichern sind. Man kann Kieselalgen zum Auftauchen bringen, indem man ein Gefäß mit Wasser und Schlamm füllt, es in schwarzes Papier einwickelt und direktes Sonnenlicht auf die Wasseroberfläche fallen lässt. Innerhalb eines Tages kommen die Kieselalgen als Schaum an die Oberfläche und können isoliert werden. ⓘ
Biochemie

| Teil einer Serie über ⓘ |
| Plankton |
|---|
 |
|
Energiequelle
Kieselalgen sind hauptsächlich photosynthetisch; einige wenige sind jedoch obligat heterotroph und können auch ohne Licht leben, sofern eine geeignete organische Kohlenstoffquelle vorhanden ist. ⓘ
Kieselsäure-Stoffwechsel
Diatomeenzellen sind in einer einzigartigen Siliziumdioxid-Zellwand enthalten, die als Kegelstumpf bekannt ist und aus zwei Klappen besteht, die Thecae genannt werden und sich normalerweise überlappen. Die biogene Kieselsäure, aus der die Zellwand besteht, wird intrazellulär durch die Polymerisation von Kieselsäuremonomeren synthetisiert. Dieses Material wird dann an die Außenseite der Zelle extrudiert und der Wand hinzugefügt. Wenn sich eine Kieselalge teilt, um zwei Tochterzellen zu erzeugen, behält bei den meisten Arten jede Zelle eine der beiden Hälften und lässt in ihr eine kleinere Hälfte wachsen. Infolgedessen wird die durchschnittliche Größe der Diatomeenzellen in der Population nach jedem Teilungszyklus kleiner. Sobald diese Zellen eine bestimmte Mindestgröße erreicht haben, teilen sie sich nicht einfach, sondern kehren diesen Rückgang um, indem sie eine Auxospore bilden. Diese vergrößert sich, um eine viel größere Zelle zu bilden, die sich dann wieder in abnehmender Größe teilt. Die Bildung von Auxosporen ist fast immer mit Meiose und sexueller Fortpflanzung verbunden. ⓘ
Der genaue Mechanismus der Übertragung der von der Diatomee aufgenommenen Kieselsäure in die Zellwand ist unbekannt. Ein Großteil der Sequenzierung von Kieselalgengen geht auf die Suche nach dem Mechanismus der Aufnahme und Ablagerung von Kieselsäure in nanoskaligen Mustern in der Frustel zurück. Die größten Erfolge in diesem Bereich wurden bei zwei Arten erzielt: Thalassiosira pseudonana, die zur Modellart wurde, da das gesamte Genom sequenziert wurde und Methoden zur genetischen Kontrolle etabliert wurden, und Cylindrotheca fusiformis, bei der die wichtigen Siliziumdioxid-Ablagerungsproteine Silaffine erstmals entdeckt wurden. Silaffine, Gruppen von polykationischen Peptiden, wurden in den Zellwänden von C. fusiformis gefunden und können komplizierte Kieselsäure-Strukturen erzeugen. Diese Strukturen wiesen Poren mit Größen auf, die für Kieselalgenmuster charakteristisch sind. Bei der Genomanalyse von T. pseudonana wurde festgestellt, dass es für einen Harnstoffzyklus kodiert, einschließlich einer höheren Anzahl von Polyaminen als in den meisten Genomen, sowie für drei verschiedene Kieseltransportgene. In einer phylogenetischen Studie über Kieselsäuretransportgene aus 8 verschiedenen Diatomeengruppen wurde festgestellt, dass der Kieselsäuretransport im Allgemeinen mit der Art zusammenhängt. In dieser Studie wurden auch strukturelle Unterschiede zwischen den Kieselsäuretransportern von pennaten (bilaterale Symmetrie) und zentrischen (radiale Symmetrie) Kieselalgen festgestellt. Die in dieser Studie verglichenen Sequenzen wurden dazu verwendet, einen vielfältigen Hintergrund zu schaffen, um Rückstände zu identifizieren, die eine unterschiedliche Funktion bei der Siliziumdioxidabscheidung haben. Darüber hinaus wurde in derselben Studie festgestellt, dass eine Reihe von Regionen innerhalb der Arten konserviert sind, was wahrscheinlich die Grundstruktur des Kieselsäuretransports darstellt. ⓘ
Diese Kieselsäuretransportproteine sind einzigartig für Diatomeen, und es wurden keine Homologe in anderen Arten, wie Schwämmen oder Reis, gefunden. Die Divergenz dieser Kieselsäuretransportgene ist auch ein Hinweis darauf, dass sich die Struktur des Proteins aus zwei sich wiederholenden Einheiten entwickelt hat, die aus fünf membrangebundenen Segmenten bestehen, was entweder auf eine Genduplikation oder eine Dimerisierung hinweist. Die Ablagerung von Kieselsäure aus dem membrangebundenen Vesikel in Diatomeen ist vermutlich auf die Aktivität von Silaffinen und langkettigen Polyaminen zurückzuführen. Dieses Silica Deposition Vesicle (SDV) wurde als ein saures Kompartiment charakterisiert, das mit Golgi-Vesikeln verschmolzen ist. Es hat sich gezeigt, dass diese beiden Proteinstrukturen in vivo Blätter aus gemustertem Siliziumdioxid mit unregelmäßigen Poren in der Größenordnung von Kieselalgenfruchtkörpern bilden. Eine Hypothese über die Art und Weise, wie diese Proteine die komplexe Struktur erzeugen, ist, dass die Reste innerhalb der SDVs konserviert sind, was leider aufgrund der begrenzten Anzahl verschiedener Sequenzen schwer zu identifizieren oder zu beobachten ist. Obwohl der genaue Mechanismus der sehr gleichmäßigen Ablagerung von Siliziumdioxid noch nicht bekannt ist, werden die Gene von Thalassiosira pseudonana, die mit Silaffinen in Verbindung stehen, als Ziele für die genetische Kontrolle der Ablagerung von Siliziumdioxid im Nanobereich angesehen. ⓘ
Harnstoff-Zyklus
Eine Besonderheit der Kieselalgen ist der Harnstoffzyklus, der sie evolutionär mit den Tieren verbindet. Das haben Andrew Allen, Chris Bowler und Kollegen in ihrer Forschung herausgefunden. Ihre 2011 veröffentlichte Erkenntnis, dass Kieselalgen über einen funktionierenden Harnstoffzyklus verfügen, war von großer Bedeutung, da man bisher davon ausging, dass der Harnstoffzyklus von den Metazoen stammt, die mehrere hundert Millionen Jahre vor den Kieselalgen auftraten. Ihre Studie zeigte, dass Diatomeen und Tiere den Harnstoffzyklus zwar zu unterschiedlichen Zwecken nutzen, dass sie aber evolutionär so miteinander verbunden sind, wie es bei Tieren und Pflanzen nicht der Fall ist. ⓘ
Pigmente
Die wichtigsten Pigmente der Kieselalgen sind die Chlorophylle a und c, Beta-Carotin, Fucoxanthin, Diatoxanthin und Diadinoxanthin. ⓘ
Speicherprodukte
Speicherprodukte sind Chrysolaminarin und Lipide. ⓘ
Taxonomie

.jpg)


Stephanodiscus hantzschii ⓘ
.jpg)
Isthmia nervosaIsthmia nervosa ⓘ

Odontella aurita ⓘ
Kieselalgen gehören zu einer großen Gruppe von Protisten, von denen viele Plastiden enthalten, die reich an Chlorophyll a und c sind. Die Gruppe wird auch als Heterokonten, Chrysophyten, Chromisten oder Stramenopile bezeichnet. Viele von ihnen sind autotroph, wie Goldalgen und Seetang, und heterotroph, wie Wasserpilze, Opaliniden und aktinophryde Heliozoen. Die Klassifizierung dieses Bereichs der Protisten ist immer noch nicht geklärt. Was die Rangordnung betrifft, so wurden sie als Abteilung, Stamm, Königreich oder etwas dazwischen behandelt. Folglich werden Kieselalgen von einer Klasse, die gewöhnlich Diatomophyceae oder Bacillariophyceae genannt wird, bis hin zu einer Abteilung (=Phylum), die gewöhnlich Bacillariophyta genannt wird, eingeordnet, wobei sich die Rangfolge der Untergruppen entsprechend ändert. ⓘ
Gattungen und Arten
Es gibt schätzungsweise 20.000 existierende Kieselalgenarten, von denen laut Guiry (2012) bisher etwa 12.000 benannt wurden (andere Quellen geben eine größere Bandbreite von Schätzungen an). Etwa 1.000-1.300 Kieselalgengattungen sind beschrieben worden, sowohl lebende als auch fossile, von denen etwa 250-300 nur als Fossilien existieren. ⓘ
Klassen und Ordnungen
Lange Zeit wurden die Kieselalgen - die entweder als Klasse (Bacillariophyceae) oder als Stamm (Bacillariophyta) behandelt wurden - in nur zwei Ordnungen unterteilt, die den zentrischen und den pennaten Kieselalgen (Centrales und Pennales) entsprechen. Diese Klassifizierung wurde 1990 von Round, Crawford und Mann grundlegend überarbeitet, die die Kieselalgen auf einer höheren Stufe (Division, entspricht dem Stamm in der zoologischen Klassifizierung) behandelten und die wichtigsten Klassifizierungseinheiten zu Klassen erhoben, Die zentrischen Kieselalgen wurden als eine einzige Klasse Coscinodiscophyceae beibehalten, während die ehemaligen pennaten Kieselalgen in zwei separate Klassen aufgeteilt wurden, die Fragilariophyceae und die Bacillariophyceae (wobei der ältere Name beibehalten, aber die Definition geändert wurde), die zusammen 45 Ordnungen umfassen, von denen die meisten neu sind. ⓘ
Heute (Mitte 2020) wird anerkannt, dass das System von Round et al. aus dem Jahr 1990 mit dem Aufkommen neuerer molekularer Arbeiten überarbeitet werden muss. Welches System es am besten ersetzen sollte, ist jedoch unklar, und die derzeit weit verbreiteten Systeme wie AlgaeBase, das World Register of Marine Species und die dazugehörige Datenbank DiatomBase sowie das System für "alles Leben" in Ruggiero et al, 2015, basieren alle auf der Behandlung von Round et al., wobei die Diatomeen als Ganzes als Klasse und nicht als Abteilung/Stamm behandelt werden und die Klassen von Round et al. auf Unterklassen reduziert werden, um eine bessere Übereinstimmung mit der Behandlung von phylogenetisch benachbarten Gruppen und den sie enthaltenden Taxa zu erreichen. (Für Referenzen siehe die einzelnen Abschnitte unten). ⓘ
Ein Vorschlag von Linda Medlin und Mitarbeitern aus dem Jahr 2004 sieht vor, einige der zentrischen Diatomeenordnungen, die als enger mit den pennaten Diatomeen verwandt gelten, als neue Klasse Mediophyceae abzuspalten, die ihrerseits enger mit den pennaten Diatomeen verwandt ist als die übrigen zentrischen Diatomeen. Diese Hypothese - später als Coscinodiscophyceae-Mediophyceae-Bacillariophyceae oder Coscinodiscophyceae+(Mediophyceae+Bacillariophyceae) (CMB)-Hypothese bezeichnet - wurde unter anderem von D.G. Mann angenommen, der sie als Grundlage für die Klassifizierung der Kieselalgen verwendet, wie sie in der Synthesereihe von Adl. et al. (2005, 2012, 2019) und auch im Bacillariophyta-Kapitel des 2017 von Archibald et al. herausgegebenen Handbook of the Protists dargestellt wird, mit einigen Änderungen, die die offensichtliche Nicht-Monophylie der ursprünglichen "Coscinodiscophyceae" von Medlin et al. widerspiegeln. Inzwischen favorisiert eine Gruppe um E.C. Theriot eine andere Hypothese der Phylogenie, die als strukturelle Gradationshypothese (SGH) bezeichnet wird und die Mediophyceae nicht als monophyletische Gruppe anerkennt, während eine andere Analyse, die von Parks et al, 2018, feststellt, dass die radial zentrischen Kieselalgen (Coscinodiscophyceae von Medlin et al.) nicht monophyletisch sind, sondern die Monophylie der Mediophyceae ohne Attheya, einer anomalen Gattung, unterstützt. Die Diskussion über die relativen Vorzüge dieser widersprüchlichen Schemata wird von den verschiedenen Beteiligten fortgesetzt. ⓘ
Adl et al. (2019)
2019 legten Adl et al. die folgende Klassifizierung der Kieselalgen vor, wobei sie anmerkten: "Diese Revision spiegelt zahlreiche Fortschritte in der Phylogenie der Diatomeen im letzten Jahrzehnt wider. Aufgrund unserer schlechten Taxonauswahl außerhalb der Mediophyceae und der pennaten Diatomeen und der bekannten und erwarteten Vielfalt aller Diatomeen erscheinen viele Kladen auf einer hohen Klassifizierungsebene (und die Klassifizierung auf höherer Ebene ist ziemlich flach)." Diese Klassifikation behandelt Kieselalgen als einen Stamm (Diatomeae/Bacillariophyta), akzeptiert die Klasse Mediophyceae von Medlin und Mitarbeitern, führt neue Subphyla und Klassen für eine Reihe ansonsten isolierter Gattungen ein und ordnet eine Reihe zuvor etablierter Taxa als Unterklassen neu ein, führt aber keine Ordnungen oder Familien auf. Aus Gründen der Klarheit wurden abgeleitete Ränge hinzugefügt (Adl. et al. verwenden keine Ränge, aber die beabsichtigten Ränge in diesem Teil der Klassifikation sind aus der Wahl der verwendeten Endungen innerhalb des verwendeten Systems der botanischen Nomenklatur ersichtlich).
- Klade Diatomista Derelle et al. 2016, emend. Cavalier-Smith 2017 (Kieselalgen und eine Untergruppe anderer Ochrophytengruppen)
- Phylum Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (Diatomeen)
- Unterstamm Leptocylindrophytina D.G. Mann in Adl et al. 2019
- Klasse Leptocylindrophyceae D.G. Mann in Adl et al. 2019 (Leptocylindrus, Tenuicylindrus)
- Klasse Corethrophyceae D.G. Mann in Adl et al. 2019 (Corethron)
- Unterstamm Ellerbeckiophytina D.G. Mann in Adl et al. 2019 (Ellerbeckia)
- Unterstamm Probosciophytina D.G. Mann in Adl et al. 2019 (Proboscia)
- Subphylum Melosirophytina D.G. Mann in Adl et al. 2019 (Aulacoseira, Melosira, Hyalodiscus, Stephanopyxis, Paralia, Endictya)
- Subphylum Coscinodiscophytina Medlin & Kaczmarska 2004, emend. (Actinoptychus, Coscinodiscus, Actinocyclus, Asteromphalus, Aulacodiscus, Stellarima)
- Subphylum Rhizosoleniophytina D.G. Mann in Adl et al. 2019 (Guinardia, Rhizosolenia, Pseudosolenia)
- Unterstamm Arachnoidiscophytina D.G. Mann in Adl et al. 2019 (Arachnoidiscus)
- Unterstamm Bacillariophytina Medlin & Kaczmarska 2004, emend.
- Klasse Mediophyceae Jouse & Proshkina-Lavrenko in Medlin & Kaczmarska 2004
- Unterklasse Chaetocerotophycidae Round & R.M. Crawford in Round et al. 1990, emend.
- Unterklasse Lithodesmiophycidae Round & R.M. Crawford in Round et al. 1990, emend.
- Unterklasse Thalassiosirophycidae Round & R.M. Crawford in Round et al. 1990
- Unterklasse Cymatosirophycidae Round & R.M. Crawford in Round et al. 1990
- Unterklasse Odontellophycidae D.G. Mann in Adl et al. 2019
- Unterklasse Chrysanthemodiscophycidae D.G. Mann in Adl et al. 2019
- Klasse Biddulphiophyceae D.G. Mann in Adl et al. 2019
- Unterklasse Biddulphiophycidae Round und R.M. Crawford in Round et al. 1990, emend.
- Biddulphiophyceae incertae sedis (Attheya)
- Klasse Bacillariophyceae Haeckel 1878, emend.
- Bacillariophyceae incertae sedis (Striatellaceae)
- Unterklasse Urneidophycidae Medlin 2016
- Unterklasse Fragilariophycidae Round in Round, Crawford & Mann 1990, emend.
- Unterklasse Bacillariophycidae D.G. Mann in Round, Crawford & Mann 1990, emend. ⓘ
Siehe Taxonomie der Kieselalgen für weitere Einzelheiten. ⓘ
Galerie
- Rasterelektronenmikroskopische Aufnahmen

Kieselalge Surirella spiralis

Kieselalge Thalassiosira sp. auf einem Membranfilter, Porengröße 0,4 μm.

Kieselalge Paralia sulcata.

Kieselalge Achanthes trinodis ⓘ



Eigenständige Zelle von Bacillaria paxillifer
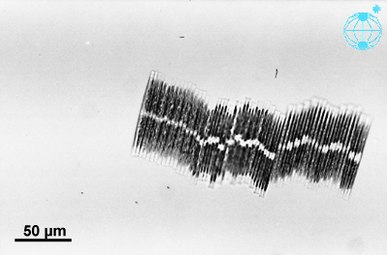
Koloniale Gruppe von Bacillaria paxillifer ⓘ

| Externes Video ⓘ | |
|---|---|
Drei Kieselalgenarten wurden zur Internationalen Raumstation geschickt, darunter die riesigen (6 mm langen) Kieselalgen der Antarktis und die einzige koloniale Kieselalge, Bacillaria paradoxa. Die Zellen von Bacillaria bewegten sich nebeneinander in teilweiser, aber entgegengesetzter Synchronität durch eine mikrofluidische Methode. ⓘ
Evolution und Fossiliennachweis
Entstehung
Die Chloroplasten von Heterokont scheinen sich von denen der Rotalgen abzuleiten und nicht wie bei den Pflanzen direkt von Prokaryoten. Dies deutet darauf hin, dass sie einen jüngeren Ursprung haben als viele andere Algen. Fossile Belege sind jedoch spärlich, und erst mit der Entwicklung der Kieselalgen selbst machen die Heterokonten einen ernsthaften Eindruck im Fossilbericht. ⓘ
Früheste Fossilien
Die frühesten bekannten fossilen Kieselalgen stammen aus dem frühen Jura (vor ca. 185 Millionen Jahren), obwohl die molekulare Uhr und die sedimentären Belege auf einen früheren Ursprung hindeuten. Es wird vermutet, dass ihr Ursprung mit dem Massenaussterben am Ende des Perms (~250 Mio. Jahre) zusammenhängt, nach dem viele Meeresnischen geöffnet wurden. Die Lücke zwischen diesem Ereignis und dem ersten Auftreten fossiler Diatomeen könnte auf eine Zeit hinweisen, in der Diatomeen nicht verkieselt waren und ihre Entwicklung kryptisch verlief. Seit dem Beginn der Verkieselung haben Kieselalgen einen bedeutenden Einfluss auf die Fossilienaufzeichnungen, wobei große fossile Ablagerungen bereits in der frühen Kreidezeit gefunden wurden und einige Gesteine, wie z. B. Kieselalgenerde, fast vollständig aus ihnen bestehen. ⓘ
Beziehung zum Siliziumzyklus
Obwohl es Kieselalgen bereits seit der Trias gibt, ist der Zeitpunkt ihres Aufstiegs und ihrer "Übernahme" des Siliziumkreislaufs jüngeren Datums. Vor dem Phanerozoikum (vor 544 Ma) wird angenommen, dass mikrobielle oder anorganische Prozesse den Siliziumkreislauf des Ozeans in geringem Maße regulierten. Danach scheint der Zyklus von Radiolarien und Kieselschwämmen dominiert (und stärker reguliert) worden zu sein, erstere als Zooplankton, letztere als sesshafte Filterfresser vor allem auf den Kontinentalschelfen. Man geht davon aus, dass der Siliziumzyklus in den letzten 100 Mio. Jahren noch stärker unter Kontrolle geraten ist, was auf die ökologische Vormachtstellung der Kieselalgen zurückzuführen ist. ⓘ
Der genaue Zeitpunkt der "Übernahme" ist jedoch nach wie vor unklar, und verschiedene Autoren interpretieren die Fossilienaufzeichnungen widersprüchlich. Einige Belege, wie die Verdrängung von Kieselschwämmen von den Schelfen, deuten darauf hin, dass diese Übernahme in der Kreidezeit (146 Ma bis 66 Ma) begann, während Belege von Radiolarien darauf hindeuten, dass die "Übernahme" erst im Känozoikum (66 Ma bis heute) begann. ⓘ
Beziehung zu Grasland
Es wird angenommen, dass die Ausbreitung von Grasland-Biomen und die evolutionäre Ausbreitung von Gräsern während des Miozäns den Fluss von löslichem Silizium in die Ozeane erhöht hat, und es wurde argumentiert, dass dies die Kieselalgen während des Känozoikums gefördert hat. Neuere Arbeiten deuten darauf hin, dass der Erfolg der Kieselalgen von der Entwicklung der Gräser entkoppelt ist, obwohl sowohl die Vielfalt der Kieselalgen als auch der Gräser ab dem mittleren Miozän stark zunahm. ⓘ
Beziehung zum Klima
Die Kieselalgenvielfalt im Känozoikum reagiert sehr empfindlich auf die globale Temperatur, insbesondere auf den Temperaturgradienten zwischen Äquator und Pol. Wärmere Ozeane, insbesondere wärmere Polarregionen, haben in der Vergangenheit nachweislich eine wesentlich geringere Diatomeenvielfalt hervorgebracht. Künftige warme Ozeane mit einer verstärkten Erwärmung der Pole, wie sie in den Szenarien für die globale Erwärmung prognostiziert wird, könnten daher theoretisch zu einem erheblichen Verlust an Kieselalgenvielfalt führen, obwohl sich nach derzeitigem Kenntnisstand nicht sagen lässt, ob dies rasch oder erst über viele Zehntausende von Jahren geschehen würde. ⓘ
Methode der Untersuchung
Die Fossilien von Kieselalgen wurden größtenteils durch die Bergung ihrer Kieselalgenfruchtkörper in marinen und nichtmarinen Sedimenten ermittelt. Obwohl Diatomeen sowohl in marinen als auch in nicht-marinen Sedimenten vorkommen, ist die Biostratigraphie der Diatomeen, die auf dem zeitlich begrenzten evolutionären Entstehen und Aussterben einzelner Taxa beruht, nur in marinen Systemen gut entwickelt und weitgehend anwendbar. Die Dauer des Vorkommens von Kieselalgenarten wurde durch die Untersuchung von Meereskernen und Gesteinsabfolgen an Land dokumentiert. Dort, wo die Biozonen der Diatomeen gut etabliert und auf die Zeitskala der geomagnetischen Polarität geeicht sind (z. B. Südpolarmeer, Nordpazifik, östlicher Äquatorialpazifik), können Altersschätzungen auf der Grundlage von Diatomeen bis auf <100 000 Jahre genau vorgenommen werden, obwohl die typische Altersauflösung für känozoische Kieselalgengemeinschaften mehrere hunderttausend Jahre beträgt. ⓘ
In Seesedimenten konservierte Diatomeen werden in großem Umfang für Paläoumweltrekonstruktionen des Klimas im Quartär verwendet, insbesondere für Seen mit geschlossenem Becken, die Schwankungen in der Wassertiefe und im Salzgehalt ausgesetzt sind. ⓘ
Isotopenaufzeichnungen

Wenn Kieselalgen sterben, können sich ihre Schalen (Frusteln) auf dem Meeresboden absetzen und zu Mikrofossilien werden. Im Laufe der Zeit werden diese Mikrofossilien als Opalablagerungen im Meeressediment begraben. Paläoklimatologie ist die Erforschung des vergangenen Klimas. Anhand von Proxydaten werden die in heutigen Sedimentproben gesammelten Elemente mit den klimatischen und ozeanischen Bedingungen in der Vergangenheit in Verbindung gebracht. Paläoklima-Proxys sind konservierte oder versteinerte physikalische Marker, die als Ersatz für direkte meteorologische oder ozeanische Messungen dienen. Ein Beispiel für Proxies ist die Verwendung von Diatomeen-Isotopenaufzeichnungen von δ13C, δ18O, δ30Si (δ13Cdiatom, δ18Odiatom, und δ30Sidiatom). Im Jahr 2015 nutzten Swann und Snelling diese Isotopenaufzeichnungen, um historische Veränderungen der Bedingungen in der photischen Zone des nordwestlichen Pazifiks zu dokumentieren, einschließlich der Nährstoffversorgung und der Effizienz der biologischen Weichteilpumpe, von der heutigen Zeit bis zum marinen Isotopenstadium 5e, das mit der letzten Zwischeneiszeit zusammenfällt. Die Spitzenwerte der Opalproduktivität im marinen Isotopenstadium stehen im Zusammenhang mit dem Zusammenbruch der regionalen Halokline-Schichtung und der erhöhten Nährstoffzufuhr in die photische Zone. ⓘ

Die anfängliche Entwicklung der Halokline und der geschichteten Wassersäule wird auf den Beginn der großen Vergletscherung der nördlichen Hemisphäre um 2,73 Ma zurückgeführt, die den Zustrom von Süßwasser in die Region durch verstärkte monsunale Regenfälle und/oder Gletscherschmelzwasser sowie die Meeresoberflächentemperaturen erhöhte. Der damit verbundene Rückgang des Auftriebs von Abyssalwasser könnte dazu beigetragen haben, dass sich weltweit kühlere Bedingungen einstellten und die Gletscher in der gesamten nördlichen Hemisphäre ab 2,73 Ma zunahmen. Während die Halokline während des späten Pliozäns und des frühen Quartärs während der Glazial-Interglazial-Zyklen vorherrschend gewesen zu sein scheint, haben andere Studien gezeigt, dass die Schichtgrenze im späten Quartär an den Enden der Glaziale und während des frühen Teils der Interglaziale aufgebrochen sein könnte. ⓘ
Diversifizierung
Die Aufzeichnungen über Kieselalgen aus der Kreidezeit sind begrenzt, aber neuere Studien zeigen eine fortschreitende Diversifizierung der Kieselalgenarten. Das Aussterbeereignis der Kreidezeit und des Paläogens, das in den Ozeanen Organismen mit Kalkskeletten dramatisch beeinträchtigte, scheint die Entwicklung der Kieselalgen relativ wenig beeinflusst zu haben. ⓘ
Umsatz
Obwohl während des Känozoikums kein Massenaussterben mariner Kieselalgen beobachtet wurde, kam es in der Nähe der Paläozän-Eozän-Grenze und an der Eozän-Oligozän-Grenze zu einem relativ raschen evolutionären Wandel der marinen Kieselalgengemeinschaften. Zwischen dem mittleren Miozän und dem späten Pliozän kam es zu verschiedenen Zeitpunkten zu einem weiteren Wandel der Artengemeinschaften als Reaktion auf die fortschreitende Abkühlung der Polarregionen und die Entwicklung endemischerer Kieselalgengemeinschaften. ⓘ
Vom Oligozän bis zum Quartär ist ein globaler Trend zu zarteren Kieselalgenfrüchten zu beobachten. Dies fällt zusammen mit einer immer stärkeren Zirkulation des Oberflächen- und Tiefenwassers der Ozeane, die durch zunehmende thermische Breitengradienten zu Beginn der großen Eisschildausdehnung in der Antarktis und die fortschreitende Abkühlung im Neogen und Quartär hin zu einer bipolar vergletscherten Welt verursacht wurde. Dies führte dazu, dass die Kieselalgen weniger Kieselsäure für die Bildung ihrer Frusteln aufnahmen. Durch die verstärkte Durchmischung der Ozeane werden Kieselsäure und andere Nährstoffe, die für das Wachstum der Kieselalgen in den Oberflächengewässern notwendig sind, erneuert, vor allem in Regionen mit küstennahem und ozeanischem Auftrieb. ⓘ
Genetik
Expressed Sequence Tagging
Im Jahr 2002 wurden die ersten Einblicke in die Eigenschaften des Genrepertoires von Phaeodactylum tricornutum anhand von 1.000 Expressed Sequence Tags (ESTs) beschrieben. In der Folge wurde die Anzahl der ESTs auf 12.000 erweitert und die Diatomeen-EST-Datenbank für funktionelle Analysen aufgebaut. Diese Sequenzen wurden verwendet, um eine vergleichende Analyse zwischen P. tricornutum und den mutmaßlich vollständigen Proteomen der Grünalge Chlamydomonas reinhardtii, der Rotalge Cyanidioschyzon merolae und der Kieselalge Thalassiosira pseudonana durchzuführen. Die Diatomeen-EST-Datenbank umfasst inzwischen über 200 000 ESTs aus Zellen von P. tricornutum (16 Bibliotheken) und T. pseudonana (7 Bibliotheken), die unter verschiedenen Bedingungen gewachsen sind, von denen viele unterschiedlichen abiotischen Belastungen entsprechen. ⓘ
Genom-Sequenzierung
Im Jahr 2004 wurde das gesamte Genom der zentrischen Kieselalge Thalassiosira pseudonana (32,4 Mb) sequenziert. 2008 folgte die Sequenzierung der pennaten Kieselalge Phaeodactylum tricornutum (27,4 Mb). Ein Vergleich der beiden Genome zeigt, dass das Genom von P. tricornutum weniger Gene enthält (10 402 gegenüber 11 776) als das von T. pseudonana; es konnte keine größere Syntenie (Genfolge) zwischen den beiden Genomen festgestellt werden. Die Gene von T. pseudonana weisen durchschnittlich 1,52 Introns pro Gen auf, während es bei P. tricornutum nur 0,79 sind, was auf eine weit verbreitete Zunahme von Introns in der zentrischen Kieselalge schließen lässt. Trotz der relativ jungen evolutionären Divergenz (90 Millionen Jahre) deutet das Ausmaß der molekularen Divergenz zwischen Zentrikern und Pennaten auf schnelle Evolutionsraten innerhalb der Bacillariophyceae im Vergleich zu anderen eukaryotischen Gruppen hin. Die vergleichende Genomik ergab auch, dass eine bestimmte Klasse von transponierbaren Elementen, die Diatomeen-Copia-ähnlichen Retrotransposons (oder CoDis), im Genom von P. tricornutum im Vergleich zu T. pseudonana signifikant vermehrt wurden und 5,8 bzw. 1 % der jeweiligen Genome ausmachen. ⓘ
Endosymbiotischer Gentransfer
Die Kieselalgengenomik lieferte viele Informationen über das Ausmaß und die Dynamik des endosymbiotischen Gentransfers (EGT). Ein Vergleich der T. pseudonana-Proteine mit Homologen in anderen Organismen ergab, dass Hunderte von ihnen ihre engsten Homologe in der Plantae-Linie haben. Der EGT-Prozess in Bezug auf Kieselalgengenome wird durch die Tatsache veranschaulicht, dass das Genom von T. pseudonana für sechs Proteine kodiert, die am engsten mit Genen verwandt sind, die vom Nukleomorph-Genom von Guillardia theta (Kryptomonade) kodiert werden. Vier dieser Gene finden sich auch in den Genomen der Rotalgenplastiden, was einen sukzessiven EGT vom Rotalgenplastid über den Rotalgenkern (Nukleomorph) zum Heterokont-Wirtskern belegt. Neuere phylogenomische Analysen von Diatomeen-Proteomen lieferten Beweise für einen Prasinophyten-ähnlichen Endosymbionten im gemeinsamen Vorfahren der Chromalveolaten, was durch die Tatsache unterstützt wird, dass 70 % der Diatomeen-Gene pflanzlichen Ursprungs aus der grünen Linie stammen und dass solche Gene auch im Genom anderer Stramenopilen zu finden sind. Daher wurde vorgeschlagen, dass Chromalveolen das Produkt einer seriellen sekundären Endosymbiose mit einer Grünalge sind, gefolgt von einer zweiten Endosymbiose mit einer Rotalge, bei der die genomischen Fußspuren der ersten erhalten blieben, das grüne Plastid jedoch verdrängt wurde. Phylogenomische Analysen der Proteome von Diatomeen und der Evolutionsgeschichte der Chromalveolen werden jedoch wahrscheinlich ergänzende genomische Daten von weniger sequenzierten Linien wie den Rotalgen nutzen. ⓘ
Horizontaler Gentransfer
Zusätzlich zum EGT kann horizontaler Gentransfer (HGT) unabhängig von einem endosymbiotischen Ereignis stattfinden. Bei der Veröffentlichung des P. tricornutum-Genoms wurde berichtet, dass mindestens 587 P. tricornutum-Gene offenbar am engsten mit bakteriellen Genen verwandt sind und mehr als 5 % des P. tricornutum-Proteoms ausmachen. Etwa die Hälfte dieser Gene ist auch im Genom von T. pseudonana zu finden, was darauf hindeutet, dass sie bereits vor langer Zeit in den Kieselalgenstamm aufgenommen wurden. ⓘ
Gentechnische Veränderungen
Um die biologischen Mechanismen zu verstehen, die der großen Bedeutung der Kieselalgen in geochemischen Kreisläufen zugrunde liegen, verwenden Wissenschaftler seit den 90er Jahren die Arten Phaeodactylum tricornutum und Thalassiosira spp. als Modellorganismen. Derzeit stehen nur wenige molekularbiologische Werkzeuge zur Verfügung, um Mutanten oder transgene Linien zu erzeugen: Plasmide, die Transgene enthalten, werden mit der biolistischen Methode oder der bakteriellen Konjugation (mit einer Ausbeute von 10-6 bzw. 10-4) in die Zellen eingebracht, und andere klassische Transfektionsmethoden wie die Elektroporation oder die Verwendung von PEG haben Berichten zufolge Ergebnisse mit geringerer Effizienz geliefert. ⓘ
Transfizierte Plasmide können entweder zufällig in die Chromosomen der Diatomee integriert werden oder als stabile zirkuläre Episomen erhalten bleiben (dank der CEN6-ARSH4-HIS3 Hefe-Zentromer-Sequenz). Das Phleomycin/Zeocin-Resistenzgen Sh Ble wird häufig als Selektionsmarker verwendet, und verschiedene Transgene wurden erfolgreich in Diatomeen eingeführt und exprimiert, wobei die Übertragung über Generationen hinweg stabil ist oder die Möglichkeit besteht, sie zu entfernen. ⓘ
Darüber hinaus ermöglichen diese Systeme nun die Verwendung des Genom-Editierwerkzeugs CRISPR-Cas, was zu einer schnellen Herstellung funktioneller Knock-out-Mutanten und einem genaueren Verständnis der zellulären Prozesse der Diatomeen führt. ⓘ
Anwendungen für den Menschen
Paläontologie
Die Zersetzung und der Zerfall von Kieselalgen führen zu organischen und anorganischen (in Form von Silikaten) Sedimenten, deren anorganische Komponente zu einer Methode zur Analyse vergangener Meeresumgebungen durch Entnahme von Bohrkernen aus dem Meeresboden oder aus dem Schlamm von Buchten führen kann, da die anorganische Substanz in die Ablagerung von Tonen und Schlämmen eingebettet ist und eine dauerhafte geologische Aufzeichnung solcher Meeresschichten bildet (siehe Kieselschlamm). ⓘ
Industriell

Kieselalgen und ihre Schalen (Frusteln) in Form von Kieselgur oder Diatomeenerde sind wichtige industrielle Rohstoffe, die zum Feinpolieren und zur Flüssigkeitsfiltration verwendet werden. Die komplexe Struktur ihrer mikroskopisch kleinen Schalen ist als Material für die Nanotechnologie vorgeschlagen worden. ⓘ
Kieselgur gilt als natürliches Nanomaterial und hat viele Verwendungszwecke und Anwendungen, wie z. B.: Herstellung verschiedener keramischer Produkte, Baukeramik, feuerfeste Keramik, spezielle Oxidkeramik, Herstellung von Materialien zur Feuchtigkeitsregulierung, Verwendung als Filtermaterial, Material in der Zementindustrie, Ausgangsmaterial für die Herstellung von Arzneimittelträgern mit verlängerter Wirkstofffreisetzung, Absorptionsmaterial im industriellen Maßstab, Herstellung poröser Keramik, Glasindustrie, Verwendung als Katalysatorträger, als Füllstoff in Kunststoffen und Farben, Reinigung von Brauchwasser, Pestizidbehälter sowie zur Verbesserung der physikalischen und chemischen Eigenschaften bestimmter Böden und andere Verwendungszwecke. ⓘ
Kieselalgen werden auch verwendet, um die Herkunft von Materialien zu bestimmen, die sie enthalten, einschließlich Meerwasser. ⓘ
Forensische
Das Hauptziel der Kieselalgenanalyse in der Gerichtsmedizin ist die Unterscheidung zwischen einem Tod durch Untertauchen und einem postmortalen Eintauchen einer Leiche in Wasser. Laboruntersuchungen können das Vorhandensein von Kieselalgen in der Leiche nachweisen. Da die auf Kieselsäure basierenden Skelette der Kieselalgen nicht leicht verwesen, können sie manchmal sogar in stark verwesten Leichen nachgewiesen werden. Da sie im Körper nicht natürlich vorkommen, kann ein Labortest, bei dem Kieselalgen der gleichen Art wie im Wasser, aus dem die Leiche geborgen wurde, nachgewiesen werden, ein guter Beweis für Ertrinken als Todesursache sein. Die Mischung der Kieselalgenarten, die in einer Leiche gefunden werden, kann die gleiche sein oder sich von der des umgebenden Wassers unterscheiden, was darauf hindeutet, ob das Opfer an dem Ort ertrunken ist, an dem die Leiche gefunden wurde. ⓘ
Nanotechnologie
Die Ablagerung von Siliziumdioxid durch Kieselalgen könnte sich auch für die Nanotechnologie als nützlich erweisen. Kieselalgenzellen stellen wiederholt und zuverlässig Ventile in verschiedenen Formen und Größen her, was es den Kieselalgen ermöglichen könnte, Strukturen im Mikro- oder Nanomaßstab herzustellen, die für eine Reihe von Geräten von Nutzen sein könnten, z. B. für optische Systeme, Halbleiter-Nanolithographie und sogar für Fahrzeuge zur Verabreichung von Medikamenten. Mit einem geeigneten künstlichen Selektionsverfahren könnten Diatomeen, die Ventile bestimmter Formen und Größen produzieren, für die Kultivierung in Chemostat-Kulturen entwickelt werden, um Komponenten im Nanomaßstab in Massenproduktion herzustellen. Es wurde auch vorgeschlagen, dass Diatomeen als Bestandteil von Solarzellen verwendet werden könnten, indem das Siliziumdioxid, das Diatomeen normalerweise zur Herstellung ihrer Zellwände verwenden, durch lichtempfindliches Titandioxid ersetzt wird. Es wurden auch Solarzellen vorgeschlagen, die aus Diatomeen Biokraftstoff herstellen. ⓘ
Geschichte der Entdeckung
Die ersten Abbildungen von Kieselalgen finden sich in einem Artikel aus dem Jahr 1703 in den Transactions of the Royal Society, der unverwechselbare Zeichnungen von Tabellaria zeigt. Obwohl die Publikation von einem ungenannten englischen Gentleman verfasst wurde, gibt es neuere Hinweise darauf, dass es sich um Charles King aus Staffordshire handelte. Erst 80 Jahre später finden wir die erste offiziell identifizierte Kieselalge, die koloniale Bacillaria paxillifera, die 1783 vom dänischen Naturforscher Otto Friedrich Müller entdeckt und beschrieben wurde. Wie viele andere nach ihm hielt er sie aufgrund ihrer Fähigkeit, sich zu bewegen, fälschlicherweise für ein Tier. Auch Charles Darwin sah auf den Kapverdischen Inseln Kieselalgenreste im Staub, obwohl er sich nicht sicher war, was sie waren. Erst später wurden sie für ihn als Kieselalgenpolygastrien identifiziert. Die Infusorien, die Darwin später in der Gesichtsfarbe der Fueguinos, der Ureinwohner Feuerlands im Süden Südamerikas, entdeckte, wurden später auf dieselbe Weise identifiziert. Noch zu seinen Lebzeiten wurde die Zugehörigkeit der Kieselalgen zu den Kieselalgengewächsen (Diatomaceae) geklärt, und Darwin rang darum, die Gründe für ihre Schönheit zu verstehen. Er tauschte sich mit dem bekannten Kryptogamisten G. H. K. Thwaites über dieses Thema aus. In der vierten Auflage von On the Origin of Species (Über die Entstehung der Arten) stellte er fest: "Wenige Objekte sind schöner als die winzigen Kieselalgengehäuse der Diatomaceae: wurden sie geschaffen, damit sie unter den hohen Kräften des Mikroskops untersucht und bewundert werden können"? und schlussfolgerte, dass ihre exquisite Morphologie einen funktionalen Hintergrund haben muss und nicht nur zum Bewundern durch den Menschen geschaffen wurde. ⓘ
Vorkommen
Kieselalgen kommen hauptsächlich im Meer und in Süßgewässern planktisch oder benthisch vor, oder sie sind auf Steinen oder Wasserpflanzen (Epiphyten) angesiedelt. Manche Arten brauchen reines und kaum verschmutztes Wasser und sind aus diesem Grunde auch Zeigerorganismen für unbelastete Gewässer. Andere Arten wiederum, die im engl. auch als agricultural guild bezeichnet werden, sind typisch für Gewässer, die durch landwirtschaftliche Einträge, bspw. durch Überdüngung, besonders belastet sind. Zu diesen werden u. a. Navicula radiosa, Melosira varians, Nitzschia palea, Diatoma vulgare oder Amphora perpusilla gezählt. Auch terrestrische Arten finden sich unter den Diatomeen; diese besiedeln Böden, in tropischen Gebieten auch Blätter von Bäumen. ⓘ
Systematik


Die Kieselalgen wurden traditionell in die radiärsymmetrischen Centrales und die bilateralsymmetrischen Pennales gegliedert. Die Centrales sind jedoch paraphyletisch, eine stabile Systematik auf molekulargenetischer Grundlage hat sich noch nicht etabliert. Nach der taxonomischen Datenbank Diatombase existiert derzeit keine allgemein akzeptierte Gliederung der Kieselalgen in Subtaxa. ⓘ
Eine weit verbreitete, aber nicht von allen Taxonomen anerkannte Gliederung könnte so aussehen, die höhere Gliederung wurde in vergleichbarer Form in die weithin verwendete Klassifikation der Eukaryoten durch Sina Adl und Kollegen übernommen. ⓘ
.jpg)
- Unterabteilung Coscinodiscophytina
- Klasse Coscinodiscophyceae
- Ordnung Asterolamprales
- Ordnung Arachnoidiscales
- Ordnung Aulacoseirales
- Gattung Aulacoseira
- Ordnung Chrysanthemodiscales
- Ordnung Corethrales
- Ordnung Coscinodiscales
- Ordnung Ethmodiscales
- Ordnung Melosirales
- Gattung Melosira
- Ordnung Orthoseirales
- Ordnung Rhizosoleniales
- Gattung Rhizosolenia
- Ordnung Stictocyclales
- Ordnung Stictodiscales
- Ordnung Leptocylindrales
- Klasse Coscinodiscophyceae
- Unterabteilung Bacillariophytina
- Klasse Mediophyceae
- Ordnung Anaulales
- Ordnung Biddulphiales
- Ordnung Chaetocerotales
- Gattung Chaetoceros
- Ordnung Cymatosirales
- Ordnung Eupodiscales (Tricertiales)
- Gattung Pleurosira
- Ordnung Hemiaulales
- Ordnung Lithodesmiales
- Ordnung Thalassiosirales
- Gattung Cyclotella
- Ordnung Toxariales
- Klasse Bacillariophyceae (entspricht weitgehend den früheren Pennales)
- Ordnung Raphoneidales
- Ordnung Rhabdonematales
- Ordnung Cocconeidales
- Gattung Algenläuse (Cocconeis)
- Ordnung Fragilariales (vermutlich paraphyletisch)
- Gattung Asterionella
- Gattung Diatoma
- Gattung Fragilaria
- Gattung Meridion
- Gattung Synedra
- Ordnung Tabellariales
- Gattung Tabellaria
- Ordnung Licmophorales
- Ordnung Thalassionematales
- Ordnung Eunotiales
- Gattung Eunotia
- Ordnung Lyrellales
- Ordnung Mastogloiales
- Ordnung Dictyoneidales
- Ordnung Cymbellales
- Gattung Cymbella
- Gattung Didymosphenia (u. a. Didymosphenia geminata)
- Gattung Gomphonema
- Gattung Rhoicosphenia
- Ordnung Achnanthales
- Gattung Achnanthes
- Ordnung Naviculales
- Gattung Diploneis
- Gattung Gyrosigma
- Gattung Navicula
- Gattung Pinnularia
- Gattung Stauroneis
- Ordnung Thalassiophysales
- Gattung Amphora
- Ordnung Bacillariales
- Gattung Bacillaria
- Gattung Nitzschia
- Ordnung Rhopalodiales
- Gattung Epithemia
- Ordnung Surirellales
- Gattung Cymatopleura
- Gattung Surirella ⓘ
- Klasse Mediophyceae
Bedeutung


Die Kieselalgen sind Hauptbestandteil des Meeresphytoplanktons und sind die Haupt-Primärproduzenten organischer Stoffe, bilden also einen wesentlichen Teil der Basis der Nahrungspyramide. Als Sauerstoff produzierende (oxygene) Phototrophe erzeugen sie auch einen großen Teil des Sauerstoffs in der Erdatmosphäre. ⓘ
Aus der relativen Arten-Zusammensetzung der Kieselalgenpopulation eines Gewässers kann recht exakt dessen Trophiegrad abgeleitet werden (Diatomeenindex), sowie weitere Gewässerparameter wie pH-Wert, Salinität, Saprobie etc. Diese Verfahren können auch auf Sedimente oder auf Öllagerstätten angewandt werden und geben dann Aufschluss über die ehemals herrschenden Lebensbedingungen. ⓘ
Zur Identifizierung der Arten wird eine Aufwuchsprobe mit Kieselalgen mit Schwefelsäure, Wasserstoffperoxid, Kaliumdichromat oder einem anderen Oxidationsmittel behandelt und so werden alle organischen Bestandteile der Probe aufgelöst. Es bleiben nur noch die reinen Siliziumdioxid-Schalen übrig. Diese werden in einem Einschlussmedium mit hohem optischem Brechungsindex (z. B. Naphrax™) eingebettet und lichtmikroskopisch mit dem Phasenkontrast-Verfahren bei ca. 1000-facher Vergrößerung identifiziert. ⓘ
Sterben die Zellen, sinken sie auf den Grund des Gewässers ab, die organischen Bestandteile werden abgebaut und die Siliziumdioxid-Schalen bilden eine Ablagerung, die sogenannte Kieselgur (Diatomeenerde). Dieser Prozess ist, insbesondere im Marinen, erst unterhalb der CCD (Calcit-Kompensationstiefe) effizient genug, um große Vorkommen zu bilden. Die entstehende Kieselgur wird in Technik und Medizin angewendet. Diatomeenschalen finden unter anderem Verwendung als Filter, zur Herstellung von Dynamit, in Zahnpasta als Putzkörper, als giftfreies Insektenbekämpfungsmittel in Form von Ungezieferpuder, sowie als reflektierendes Material in der Farbe, die für Fahrbahnmarkierungen im Straßenbau verwendet wird. Außerdem finden Kieselalgen in der forensischen Medizin Verwendung (Diatomeennachweis). Ihre Aussagekraft für den Nachweis eines Ertrinkungstodes wird jedoch kontrovers diskutiert. ⓘ
Insbesondere im 19. Jahrhundert dienten Diatomeen zur Anfertigung ästhetischer mikroskopischer Präparate, deren gemeinsame Betrachtung in Salons ein beliebter Zeitvertreib in höheren gesellschaftlichen Kreisen war. Zur höchsten Perfektion in der handwerklich extrem schwierigen Anfertigung solcher Präparate hat es Johann Diedrich Möller aus Wedel bei Hamburg gebracht. ⓘ
Marine Diatomeen können zu Vergiftungen bei Mensch und Tier führen, da einige Arten, insbesondere Pseudo-nitzschia, Nitzschia oder Amphora Domoinsäure produzieren. In filtirierende Meerwasser-Organismen, wie z. B. Muscheln, oder in sich von diesen Tieren ernährende Arten, wie z. B. Fischen, können sich diese Diatomeen akkumulieren. Kommt es zum Verzehr solcher mit Domoinsäure angereicherten Organismen durch den Menschen, treten Vergiftungserscheinungen auf, die als Amnesic Shellfish Poisoning (ASP) bezeichnet werden. Als Symptome treten insbesondere Gedächtnisverlust, Übelkeit, Krämpfe, Durchfall, Kopfschmerz und Atembeschwerden auf. ⓘ