Stachelhäuter
| Stachelhäuter | |
|---|---|

| |
| Ausgestorbene Stachelhäuter der fünf Klassen: Protoreaster linckii (Asteroidea), Ophiocoma scolopendrina (Ophiuroidea), Stomopneustes variolaris (Echinoidea), Oxycomanthus bennetti (Crinoidea), Actinopyga echinites (Holothuroidea) | |
| Wissenschaftliche Klassifizierung | |
| Königreich: | Tierreich (Animalia) |
| Unterlebewesen: | Eumetazoa |
| Klade: | Parahoxozoa |
| Klade: | Bilateria |
| Klade: | Nacktwassertiere |
| Überstamm: | Deuterostomia |
| Klade: | Ambulacraria |
| Stamm: | Stachelhäuter (Echinodermata) Bruguière, 1791 [ex Klein, 1734] |
| Typusgattung | |
| Echinus Linnaeus, 1758
| |
| Subphyla und Klassen | |
|
Homalozoa † Gill & Caster, 1960
Crinozoa
Asterozoa
Stachelhäuter (Echinozoa)
Blastozoa †
†=Ausgestorben | |
Ein Echinoderm (/ɪˈkaɪnəˌdɜːrm, ˈɛkə-/) ist ein Mitglied des Stammes Echinodermata (/ɪˌkaɪnoʊˈdɜːrmətə/). Die erwachsenen Tiere sind an ihrer (in der Regel fünfzackigen) radialen Symmetrie zu erkennen und gehören zu den Seesternen, Schlangensternen, Seeigeln, Sanddollars und Seegurken sowie zu den Seelilien oder "Steinlilien". Ausgewachsene Stachelhäuter findet man auf dem Meeresboden in allen Meerestiefen, von der Gezeitenzone bis zur Abyssalzone. Der Stamm umfasst etwa 7.000 lebende Arten und ist damit nach den Chordatieren die zweitgrößte Gruppe der Deuterostomier. Die Stachelhäuter sind der größte rein marine Stamm. Die ersten endgültigen Stachelhäuter erschienen zu Beginn des Kambriums. ⓘ
Die Stachelhäuter sind sowohl aus ökologischer als auch aus geologischer Sicht von Bedeutung. Ökologisch gesehen gibt es nur wenige andere Gruppen, die in der biotischen Wüste der Tiefsee und auch in flacheren Meeren so zahlreich vorkommen. Die meisten Stachelhäuter sind in der Lage, sich ungeschlechtlich fortzupflanzen und Gewebe, Organe und Gliedmaßen zu regenerieren; in einigen Fällen können sie eine vollständige Regeneration aus einer einzigen Gliedmaße vornehmen. Geologisch gesehen liegt der Wert der Stachelhäuter in ihren verknöcherten Skeletten, die einen wichtigen Beitrag zu vielen Kalksteinformationen leisten und wertvolle Hinweise auf die geologische Umgebung liefern können. Im 19. und 20. Jahrhundert wurden sie in der Regenerationsforschung am häufigsten verwendet. Einige Wissenschaftler sind der Ansicht, dass die Ausbreitung der Stachelhäuter für die mesozoische Meeresrevolution verantwortlich war. ⓘ
| Stachelhäuter ⓘ | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Verschiedene Stachelhäuter, oben links Seesterne, oben rechts Haarsterne, mitte rechts ein Seeigel, unten rechts Seewalzen und unten links Schlangensterne. | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Echinodermata | ||||||||||||
| Bruguière, 1791 [ex Klein, 1734] |
Die Stachelhäuter (Echinodermata) (von griech. ἐχῖνος echinos „Igel“ und δέρμα derma „Haut“) sind ein zu den Deuterostomiern gehörender Tierstamm. Weltweit sind etwa 6300 rezente Arten der Stachelhäuter bekannt, womit sie die zweitgrößte Tiergruppe innerhalb der Neumundtiere (Deuterostomia) nach den Chordatieren (Chordata) bilden. ⓘ
Es handelt sich bei ihnen durchweg um Meeresbewohner, die bis auf wenige Tiefseearten reine Bodenbewohner sind – d. h. vorwiegend im Flachmeer leben. Neben ihrer weiten Verbreitung sind sie auch geologisch von Bedeutung, weil ihre abgestorbenen Schalen und Skelettteile wesentlich zur Bildung von Kalkgesteinen beitragen. Bei Vorherrschen einer Skelettart erhalten sie Namen wie Trochiten- oder Crinoidenkalk, die sich unter anderem in den Kalkalpen finden. Mengenmäßig überwiegen aber die Ablagerungen der winzigen, schwebenden Kalkalgen. ⓘ
Die Echinodermata werden traditionell in die meist gestielten Pelmatozoa sowie die freilebenden Eleutherozoa unterteilt. Zu den Pelmatozoa zählen mehrere als „Seelilien“ zusammengefasste Taxa sowie die sekundär freilebenden Haarsterne (Crinoidea). Die Eleutherozoa umfassen die bekannteren Gruppen der Stachelhäuter: die Seesterne (Asteroidea) und die Schlangensterne (Ophiuroidea), die Seeigel (Echinoidea) sowie die Seewalzen (Holothuroidea). Die seit 1986 als eine weitere Klasse identifizierten Seegänseblümchen (Concentricycloidea) werden inzwischen zu den Seesternen gestellt. ⓘ
Taxonomie und Evolution
Der Name Stachelhäuter stammt aus dem Altgriechischen ἐχῖνος (ekhînos) "Igel" und δέρμα (dérma) "Haut"). Stachelhäuter sind Bilaterer, das heißt, ihre Vorfahren waren spiegelsymmetrisch. Unter den Bilateralen gehören sie zur Abteilung der Deuterostomier, was bedeutet, dass die Blastopore, die erste Öffnung, die sich während der Embryonalentwicklung bildet, zum Anus und nicht zum Mund wird. Charakteristisch für erwachsene Stachelhäuter sind ein Wassergefäßsystem mit äußeren Röhrenfüßen und ein kalkhaltiges Endoskelett, das aus Gehörknöchelchen besteht, die durch ein Netz aus Kollagenfasern verbunden sind. ⓘ
Phylogenie
In der Vergangenheit glaubten die Taxonomen, dass die Ophiuroidea mit den Asteroidea oder mit den (Holothuroidea + Echinoidea) verwandt sind. Eine 2014 durchgeführte Analyse von 219 Genen aus allen Klassen von Stachelhäutern revidierte jedoch den phylogenetischen Baum. Eine unabhängige Analyse der RNA-Transkriptome von 23 Arten aus allen Klassen von Stachelhäutern im Jahr 2015 ergab denselben Baum. ⓘ
- Externe Phylogenie ⓘ
Der Kontext der Stachelhäuter innerhalb der Bilateria ist:
- Interne Phylogenie ⓘ
Vielfalt
Es gibt etwa 7.000 lebende Arten von Stachelhäutern sowie etwa 13.000 ausgestorbene Arten. Alle Stachelhäuter gehören zu den Meerestieren, aber sie kommen in Lebensräumen vor, die von flachen Gezeitenzonen bis zu abgrundtiefen Gewässern reichen. Traditionell werden zwei Hauptuntergruppen unterschieden: die bekannteren beweglichen Stachelhäuter (Eleutherozoa), zu denen die Seesterne (Asteroidea) mit etwa 1.745 Arten, die Schlangensterne (Ophiuroidea) mit etwa 2.300 Arten, die Seeigel (Echinoidea) mit etwa 900 Arten und die Seegurken (Holothuroidea) mit etwa 1.430 Arten gehören, und die beweglichen Stachelhäuter (Pelmatozoa), von denen einige sessil sind, während andere beweglich sind. Diese bestehen aus den Crinoidea (Federsterne und Seelilien mit etwa 580 Arten) und den ausgestorbenen Blastoiden und Paracrinoiden. ⓘ
- Die Untergliederung der Stachelhäuter

Ein Schlangenstern, Ophionereis reticulata

Eine Seegurke aus Malaysia

Seesterne weisen eine breite Palette von Farben auf

Ein Seeigel, Strongylocentrotus purpuratus

Seelilie auf einem Korallenriff ⓘ




Fossile Geschichte
Das älteste bekannte Stachelhäuterfossil ist Arkarua aus dem Präkambrium von Australien. Diese Fossilien sind scheibenförmig, mit radialen Rippen am Rand und einer fünfzackigen zentralen Vertiefung, die mit radialen Linien versehen ist. Die Fossilien weisen jedoch kein Stereom oder eine innere Struktur auf, die auf ein Wassergefäßsystem hinweist, so dass sie nicht eindeutig identifiziert werden können. ⓘ
Die ersten allgemein anerkannten Stachelhäuter tauchen im Unterkambrium auf; die Asterozoen erschienen im Ordovizium, während die Seelilien im Paläozoikum eine dominierende Gruppe waren. Stachelhäuter haben ein umfangreiches Fossilarchiv hinterlassen. Es wird angenommen, dass der Vorfahre aller Stachelhäuter ein einfaches, bewegliches, zweiseitig symmetrisches Tier mit Mund, Darm und Anus war. Dieser Vorläuferorganismus lebte in einer anhängenden Lebensform mit Suspensionsnahrung und entwickelte eine radiale Symmetrie. Dennoch sind die Larven aller Stachelhäuter zweiseitig symmetrisch, und alle entwickeln bei der Metamorphose eine radiale Symmetrie. Wie ihre Vorfahren klammern sich die Seesterne und Seelilien auch bei der Verwandlung in ihre erwachsene Form noch an den Meeresboden. ⓘ
Die ersten Stachelhäuter waren unbeweglich, entwickelten sich aber zu Tieren, die sich frei bewegen konnten. Sie entwickelten bald Endoskelettplatten mit Stereomstruktur und äußere Zilienrillen zur Nahrungsaufnahme. Die paläozoischen Stachelhäuter waren kugelförmig, mit dem Substrat verbunden und mit der Mundfläche nach oben gerichtet. Diese frühen Stachelhäuter hatten seitlich am Körper verlaufende Ambulakralrillen, die auf beiden Seiten von Brachiolen gesäumt waren, ähnlich den Fiederblättern der modernen Seelilien. Schließlich kehrten alle Stachelhäuter mit Ausnahme der Seelilien ihre Ausrichtung um, so dass der Mund nach unten zeigte. Bevor dies geschah, hatten die Podien wahrscheinlich eine Nahrungsfunktion, so wie sie heute bei den Seelilien zu finden sind. Die Fortbewegungsfunktion der Podien kam erst später hinzu, als die Podien durch die Neuausrichtung des Mundes zum ersten Mal in Kontakt mit dem Substrat kamen. ⓘ
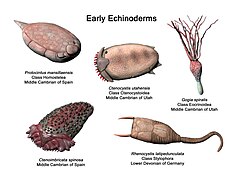
Frühe Stachelhäuter Ctenoimbricata, Ctenocystis, Gogia, Protocintus und Rhenocystis

Der ordovizische Zystoide Echinosphaerites aus dem nordöstlichen Estland

Fossile Kronen von Seelilien




Erste sichere Funde von Stachelhäutern sind bereits seit dem frühen Kambrium von über 540 Millionen Jahren bekannt. Allerdings gibt es auch Fossilien aus dem späten Präkambrium (Arkarua, Tribrachidium), die häufig dieser Tiergruppe zugeordnet werden. Innerhalb einer ersten Radiation im Kambrium entstanden mehrere heute nicht mehr existierende Großgruppen. Die Helicoplacoidea, spiralig aufgebaute Tiere, starben bereits im frühen Kambrium wieder aus. Erfolgreicher sind die asymmetrischen Carpoidea, die in mehreren wahrscheinlich nicht näher verwandten Taxa existierten. Von den vier Taxa überdauerten zwei das Kambrium, auch diese verschwinden jedoch im Laufe des Paläozoikums. Nicht eingeordnet wurden bislang die Vertreter der Gattungen Peridionites und Cymbionites, die während des mittleren Kambriums lebten. ⓘ
Mit den wenig erfolgreichen Gruppen des Kambriums entstanden auch die bereits fünfstrahligen Eocrinoidea und Edrioasteroidea, aus denen sich die modernen Vertreter der Stachelhäuter ableiten lassen. Diese stellen die Basis für eine weitere Radiation der Stachelhäuter im unteren Ordovizium dar. Die Eocrinoidea zeichneten sich durch eine aufwärts gerichtete Mundöffnung sowie den Erwerb von Körperanhängen in Form von Brachiolen (selten echten Armen) aus. Innerhalb dieser Gruppe entwickelten sich die Cystoidea und die Blastoidea sowie die Parablastoidea und die Paracrinoidea. Ob die heute noch existierenden Seelilien und Haarsterne ebenfalls auf diese Gruppe zurückgehen, ist nicht bekannt. ⓘ
Die Edrioasteroidea besaßen keine Verankerungsorgane und auch keine Brachiolen, bei ihnen kam es allerdings zu einer Umorientierung der Mundöffnung auf die Unterseite der Tiere, während der After auf die Oberseite wanderte. Diese Tiere bildeten die Wurzel der Eleutherozoa, zu denen auch die heute noch lebenden Seesterne, Schlangensterne, Seeigel und Seewalzen gehören. ⓘ
Anatomie und Physiologie
Stachelhäuter haben sich aus Tieren mit bilateraler Symmetrie entwickelt. Obwohl erwachsene Stachelhäuter eine pentaradiale Symmetrie aufweisen, sind ihre Larven flimmernde, frei schwimmende Organismen mit bilateraler Symmetrie. Später, während der Metamorphose, wächst die linke Seite des Körpers auf Kosten der rechten, die schließlich absorbiert wird. Die linke Seite wächst dann in einer pentaradialen Symmetrie, bei der der Körper in fünf Teilen um eine zentrale Achse angeordnet ist. Innerhalb der Asterozoa kann es einige Ausnahmen von dieser Regel geben. Die meisten Seesterne der Gattung Leptasterias haben sechs Arme, obwohl auch fünfarmige Exemplare vorkommen können. Auch bei den Brisingida gibt es einige sechsarmige Arten. Unter den Schlangensternen gibt es sechsarmige Arten wie Ophiothela danae, Ophiactis savignyi und Ophionotus hexactis, und Ophiacantha vivipara hat oft mehr als sechs Arme. ⓘ
Stachelhäuter weisen in einem bestimmten Lebensstadium in Teilen ihres Körpers eine sekundäre Radialsymmetrie auf, wahrscheinlich eine Anpassung an eine sessile oder sich langsam bewegende Existenz. Viele Seelilien und einige Seesterne sind in Vielfachen der Grundzahl fünf symmetrisch; Seesterne wie Labidiaster annulatus besitzen bis zu fünfzig Arme, während die Seelilie Comaster schlegelii zweihundert hat. ⓘ
Haut und Skelett

Auch wenn der Name Stachelhäuter es anders vermuten lässt, haben die Tiere kein Außenskelett. Die äußerste Schicht wird vielmehr durch eine dünne und einschichtige Epidermis gebildet, die vor allem aus wenig differenzierten Stützzellen mit jeweils einer Cilie besteht. Dazwischen liegen Drüsenzellen, Mechanorezeptoren und Pigmentzellen. Die Drüsenzellen produzieren dabei Schleim und Klebesekrete und sie kommen entsprechend besonders häufig im Bereich der Saugfüßchen bei Seeigeln, Seesternen und Seewalzen sowie an den Mundtentakeln der Seewalzen vor. Bei einigen Seeigeln, vor allem den Vertretern der Toxopneustidae, können die Drüsenzellen auch Giftstoffe enthalten, die auch beim Menschen als Neurotoxin wirken und gefährlich sind. ⓘ
Die Pigmentzellen sind für die Färbung der Tiere verantwortlich. Sie können auf Licht reagieren, sodass es bei vielen Arten zu unterschiedlichen Tages- und Nachtzeichnungen kommt. Diese Reaktion kann sehr schnell erfolgen – so verfärbt sich der Seeigel Centrostephanus longispinus in nur 50 Minuten von einem Graubraun in ein Tiefschwarz, wenn er plötzlich Licht ausgesetzt wird. Die Farben selbst kommen durch eine Mischung verschiedener Farbstoffe zustande, darunter die dunklen Melanine, rote Carotinoide und Carotinproteine, die blau, grün oder violett sein können. Hinzu kommen Naphtochinone und bei den Diademseeigeln auch noch physikalische Effekte durch reflektierende Iridophoren. ⓘ

Das Skelett liegt unterhalb der Epidermis im Mesoderm und besteht aus Calcit mit einem Magnesiumoxidanteil von drei bis fünfzehn Prozent. Da Magnesium-Ionen bei gleicher Ladung kleiner sind als die im Calcit vorherrschenden Calcium-Ionen, bewirken sie eine Erhöhung der Packungsdichte der Calcit-Kristalle und eine Verstärkung der chemischen Bindungen, was wiederum eine Steigerung der Härte und Festigkeit des Materials zur Folge hat. Dieser Effekt kommt insbesondere bei den Zähnen der Stachelhäuter zum Tragen. Durch seine periphere Lage bestimmt das Skelett maßgeblich die Gestalt der Tiere. Das Kalkskelett entsteht dabei nicht durch einfache Abscheidung von Epidermiszellen, sondern wird innerhalb eines Zellverbandes von Skelettbildungszellen (Sclerocyten) angelegt. Das entstehende Gebilde, Stereom genannt, ist stark porös, wodurch eine zugleich leichte aber auch feste Konstruktion entsteht. Das Stereom verwächst zu Platten, die als Ossikel bezeichnet werden. Das Wachstum kann in alle Richtungen erfolgen, sodass nach dem Verlust eines Körperteils dieser wieder ersetzt werden kann. Größere Elemente bilden Gelenke im Skelett aus, die durch Muskulatur beweglich werden. ⓘ
Auch die Stacheln der Seesterne und Seeigel sind mit dem Skelett gelenkig verbunden und können gegen die darunterliegenden Skelettplatten bewegt werden. Die Stacheln selbst sind spezielle Bildungen des Stereoms, die nur aus einer hohl zusammengerollten Skelettplatte bestehen. Wie alle anderen Skelettelemente sind auch sie von Epidermis überzogen. Eine Sonderbildung des Skeletts bei den Seeigeln und Seesternen stellen die pinzettenartigen Pedicellarien dar, die sich wahrscheinlich aus Stacheln entwickelt haben. Sie bestehen aus mehreren Elementen und können zur Abwehr von Parasiten und sich festsetzenden Larven anderer Meerestiere sowie zum Beutefang eingesetzt werden. Weitere Skelettelemente sind der Kieferapparat der Seeigel (Laterne des Aristoteles) aus 40 einzelnen Skelettplatten und der Kalkring der Seewalzen, der die Mundöffnung stabilisiert. Bei den Seewalzen ist die Epidermis außerdem mit mikroskopisch kleinen Kalknadeln (Spiculae) gespickt. Auch der Stiel der Seelilien besteht aus einzelnen Skelettelementen, den scheibenförmigen Trochiten. ⓘ
Stachelhäuter haben ein mesodermales Skelett in der Lederhaut, das aus Platten auf Kalzitbasis, den so genannten Ossikeln, besteht. Wären sie fest, würden sie ein schweres Skelett bilden, daher haben sie eine schwammartige poröse Struktur, die als Stereom bezeichnet wird. Die Gehörknöchelchen können miteinander verschmolzen sein, wie bei den Seeigeln, oder sie sind gelenkig und bilden flexible Gelenke, wie bei den Armen von Seesternen, Schlangensternen und Seelilien. Die Gehörknöchelchen können äußere Vorsprünge in Form von Stacheln, Körnchen oder Warzen aufweisen und werden von einer zähen Epidermis getragen. Skelettelemente werden manchmal in spezialisierter Weise eingesetzt, wie z. B. das als "Aristoteleslaterne" bezeichnete Kauorgan der Seeigel, die stützenden Stängel der Seelilien und der strukturelle "Kalkring" der Seegurken. ⓘ
Obwohl einzelne Gehörknöchelchen robust sind und sich leicht versteinern lassen, sind vollständige Skelette von Seesternen, Schlangensternen und Seelilien im Fossilbericht selten. Andererseits sind Seeigel in Kreide- oder Kalksteinschichten oft gut erhalten. Während der Fossilisierung werden die Hohlräume im Stereom mit Kalzit ausgefüllt, das mit dem umgebenden Gestein zusammenhängt. Beim Aufbrechen eines solchen Gesteins können Paläontologen ausgeprägte Spaltmuster und manchmal sogar die komplizierte innere und äußere Struktur der Probe beobachten. ⓘ
Eine Besonderheit der meisten Stachelhäuter ist eine besondere Art von Gewebe, das so genannte Fangbindegewebe. Dieses auf Kollagen basierende Material kann seine mechanischen Eigenschaften unter nervöser Kontrolle und nicht durch Muskelkraft verändern. Dank dieses Gewebes kann sich ein Seestern nicht nur flexibel auf dem Meeresboden bewegen, sondern auch starr werden, wenn er eine Muschel aufbricht oder sich selbst daran hindert, aus einer Spalte herausgezogen zu werden. In ähnlicher Weise können Seeigel ihre normalerweise beweglichen Stacheln als Verteidigungsmechanismus aufrecht halten, wenn sie angegriffen werden. ⓘ
Das Wassergefäßsystem

Stachelhäuter verfügen über ein einzigartiges Wassergefäßsystem, ein Netz von flüssigkeitsgefüllten Kanälen, die vom Coelom (Körperhöhle) abgeleitet sind und dem Gasaustausch, der Ernährung, der sensorischen Wahrnehmung und der Fortbewegung dienen. Dieses System variiert zwischen den verschiedenen Stachelhäuterklassen, öffnet sich aber in der Regel durch ein siebartiges Madreporit an der aboralen (oberen) Oberfläche des Tieres nach außen. Das Madreporit ist mit einem schmalen Kanal, dem Steinkanal, verbunden, der in einen Ringkanal übergeht, der den Mund oder die Speiseröhre umgibt. Der Ringkanal verzweigt sich in eine Reihe von Radialkanälen, die bei Asteroiden entlang der Arme verlaufen und bei Echinoiden in den Ambulakralbereichen an den Test anschließen. Von den Radialkanälen zweigen kurze Seitenkanäle ab, die jeweils in einer Ampulle enden. Ein Teil der Ampulle kann durch eine Pore (oder ein Porenpaar bei Seeigeln) nach außen ragen und ein Podium oder einen Röhrenfuß bilden. Das Wassergefäßsystem trägt zur Verteilung der Nährstoffe im Körper des Tieres bei; am deutlichsten ist es an den Röhrenfüßen zu erkennen, die sich durch die Umverteilung von Flüssigkeit zwischen dem Fuß und der inneren Ampulle ausdehnen oder zusammenziehen können. ⓘ
Die Organisation des Wassergefäßsystems ist bei den Ophiuroiden etwas anders, wo sich das Madreporit auf der Mundoberfläche befinden kann und die Podien keine Saugnäpfe haben. Bei den Holothuroiden ist das System reduziert, oft mit wenigen Röhrenfüßen außer den spezialisierten Ernährungstentakeln, und das Madreporit öffnet sich zum Coelom hin. Einige Holothuroiden wie die Apodida haben keine Röhrenfüße und keine Kanäle entlang des Körpers; andere haben Längskanäle. Die Anordnung der Röhrenfüße ist ähnlich wie bei den Asteroiden, aber die Röhrenfüße haben keine Saugnäpfe und werden in einer Hin- und Herbewegung benutzt, um die von den Armen eingefangenen Nahrungspartikel zum zentralen Mund zu befördern. Bei den Asteroiden wird die gleiche Bewegung eingesetzt, um das Tier über den Boden zu bewegen. ⓘ
Einzigartig ist das innere Kanalsystem (Ambulacralsystem), dessen äußere Fortsätze als Saugfüßchen der Fortbewegung der Tiere oder als Tentakel dem Nahrungserwerb dienen. Es hat sich in der Ontogenese vor allem aus dem Hydrocoel entwickelt und besteht aus einem Ringkanal um den Darm sowie die in die einzelnen Radien führenden Ambulacral- oder Radialkanäle. Auch die Madreporenplatte ist über einen verhärteten Kanal, den Steinkanal, mit dem System verbunden und stellt (außer bei einigen Seewalzen) die Verbindung zum freien Wasser dar. ⓘ
An den Radialkanälen liegt der Füßchen- und Tentakelapparat der Tiere. Dabei handelt es sich um Fortsätze, die rechts und links vom Radiärkanal angeordnet sind und mit diesem durch einen dünnen Zuleitungskanal in Verbindung stehen. Sie können durch den Kalkpanzer der Tiere nach außen geführt werden und dienen hier dem Nahrungserwerb, dem Gasaustausch, der Fortbewegung, der Exkretion sowie der Osmoregulation. Sie sind bei den verschiedenen Gruppen der Stachelhäuter unterschiedlich aufgebaut. So liegen sie bei den Seelilien und Haarsternen vor allem an den Transportrinnen für die Nahrungsaufnahme, den Pinnulae, und transportieren die Nahrungspartikel zur Mundöffnung. Bei den Seesternen, Seeigeln und Seewalzen bilden sie Füßchen mit einer muskulösen inneren Ampulle, die die Bewegung durch eine Regulation des Flüssigkeitsdrucks steuert. ⓘ
Andere Organe
Stachelhäuter besitzen ein einfaches Verdauungssystem, das je nach Ernährungsweise der Tiere variiert. Seesterne sind meist Fleischfresser und haben einen Mund, eine Speiseröhre, einen zweiteiligen Magen, einen Darm und einen Enddarm, wobei sich der Anus in der Mitte der aboralen Körperoberfläche befindet. Mit wenigen Ausnahmen besitzen die Mitglieder der Ordnung Paxillosida keinen Anus. Bei vielen Seesternarten kann der große Herzmagen umgedreht werden, um Nahrung außerhalb des Körpers zu verdauen. Einige andere Arten sind in der Lage, ganze Nahrungsmittel, wie z. B. Mollusken, aufzunehmen. Schlangensterne, die sich unterschiedlich ernähren, haben einen blinden Darm ohne Darm und Anus; sie scheiden Nahrungsreste durch den Mund aus. Seeigel sind Pflanzenfresser und benutzen ihre spezialisierten Mundwerkzeuge zum Abweiden, Zerreißen und Kauen ihrer Nahrung, hauptsächlich Algen. Sie haben eine Speiseröhre, einen großen Magen und einen Enddarm mit dem Anus an der Spitze des Testes. Seegurken sind hauptsächlich Detritivoren, die das Sediment mit modifizierten Röhrenfüßen um ihren Mund, den Bukkal-Tentakeln, durchwühlen. Sand und Schlamm begleiten ihre Nahrung durch ihren einfachen Darm, der aus einem langen gewundenen Darm und einer großen Kloake besteht. Stachelhäuter sind Suspensionsfresser und fangen passiv Plankton, das in ihre ausgestreckten Arme driftet. Die in Schleim eingeschlossenen Nahrungsballen werden in den Mund befördert, der durch eine Schleife, die aus einer kurzen Speiseröhre und einem längeren Darm besteht, mit dem Anus verbunden ist. ⓘ
Die Coelomhöhlen von Stachelhäutern sind komplex. Neben dem Wassergefäßsystem verfügen Stachelhäuter über ein hämales Coelom, ein periviszerales Coelom, ein gonadales Coelom und häufig auch ein perihämales Coelom. Während der Entwicklung ist das Coelom der Stachelhäuter in das Metacoel, Mesocoel und Protocoel (auch Somatocoel, Hydrocoel bzw. Axocoel genannt) unterteilt. Das Wassergefäßsystem, das Hämalsystem und das Perihämalsystem bilden das röhrenförmige Coelomsystem. Eine Besonderheit der Stachelhäuter ist die Tatsache, dass sie sowohl über ein coelomisches Kreislaufsystem (das Wassergefäßsystem) als auch über ein hämisches Kreislaufsystem verfügen, während die meisten anderen Tiergruppen nur eines der beiden Systeme besitzen. ⓘ
Hämale und perihämale Systeme sind vom ursprünglichen Coelom abgeleitet und bilden ein offenes und reduziertes Kreislaufsystem. Dieser besteht in der Regel aus einem zentralen Ring und fünf radialen Gefäßen. Es gibt kein echtes Herz, und dem Blut fehlt oft jegliches Atmungspigment. Der Gasaustausch erfolgt über dermale Branchiae oder Papulae bei Seesternen, Genitalschleimbeutel bei Schlangensternen, peristomale Kiemen bei Seeigeln und Kloakenbäume bei Seegurken. Der Gasaustausch findet auch über die Röhrenfüße statt. Da Stachelhäuter keine spezialisierten Ausscheidungsorgane besitzen, diffundieren stickstoffhaltige Abfälle, vor allem in Form von Ammoniak, über die Atmungsflächen nach außen. ⓘ
Die Coelomflüssigkeit enthält die Coelomozyten, die Immunzellen. Es gibt mehrere Arten von Immunzellen, die sich je nach Klasse und Art unterscheiden. Alle Klassen verfügen über eine Art phagozytischer Amöbozyten, die eingedrungene Partikel und infizierte Zellen verschlingen, aggregieren oder verklumpen und an der Zytotoxizität beteiligt sein können. Diese Zellen sind in der Regel groß und körnig und gelten als wichtigste Verteidigungslinie gegen potenzielle Krankheitserreger. Je nach Klasse verfügen Stachelhäuter über Kugelzellen (für Zytotoxizität, Entzündung und antibakterielle Aktivität), Vibrationszellen (für die Bewegung der Coelomflüssigkeit und die Gerinnung) und Kristallzellen (die bei Seegurken der Osmoregulation dienen können). Die Coelomozyten scheiden antimikrobielle Peptide gegen Bakterien aus und verfügen über eine Reihe von Lektinen und Komplementproteinen als Teil eines angeborenen Immunsystems, das noch charakterisiert werden muss. ⓘ
Stachelhäuter haben ein einfaches radiales Nervensystem, das aus einem modifizierten Nervennetz miteinander verbundener Neuronen besteht und kein zentrales Gehirn besitzt, obwohl einige von ihnen Ganglien besitzen. Die Nerven strahlen von zentralen Ringen um den Mund in jeden Arm oder entlang der Körperwand aus; die Verzweigungen dieser Nerven koordinieren die Bewegungen des Organismus und die Synchronisation der Röhrenfüße. Seesterne haben Sinneszellen im Epithel und verfügen über einfache Augenpunkte und berührungsempfindliche, tentakelartige Röhrenfüße an den Spitzen ihrer Arme. Seeigel haben keine besonderen Sinnesorgane, verfügen aber über Statozysten, die ihnen bei der Orientierung durch die Schwerkraft helfen, und haben Sinneszellen in ihrer Epidermis, insbesondere in den Röhrenfüßen, Stacheln und Pedicellariae. Schlangensterne, Seelilien und Seegurken haben im Allgemeinen keine Sinnesorgane, aber einige wühlende Seegurken der Ordnung Apodida haben eine einzelne Statozyste neben jedem Radialnerv, und einige haben einen Augenfleck an der Basis jedes Tentakels. ⓘ
Bei Seeigeln und Seegurken nehmen die Keimdrüsen zumindest zeitweise einen Großteil der Körperhöhlen ein, während die weniger voluminösen Seelilien, Schlangensterne und Seesterne zwei Keimdrüsen in jedem Arm haben. Während man annimmt, dass die Vorfahren der modernen Stachelhäuter eine einzige Genitalöffnung hatten, besitzen viele Organismen mehrere Gonoporen, durch die Eier oder Spermien freigesetzt werden können. ⓘ

Das Blutgefäßsystem oder besser Hämalsystem besteht aus einem zentralen Ring (Hämalring) mit fünf Kanälen entlang der Ambulacralkanäle. Ein weiterer Kanal zieht zum Axialorgan, welches die Verbindungsstelle zwischen dem Hämalsystem und dem Ambulacralsystem darstellt und als kontraktives Organ die Flüssigkeitszirkulation regelt. Die Exkretion erfolgt über das Ambulacralsystem. ⓘ
Die Gonaden füllen vor allem bei den Seeigeln und Seewalzen die gesamte Leibeshöhle aus, bei den Seelilien und Seesternen liegen sie in den Armen, bei den Seesternen jeweils zwei aufgefächerte Gonaden pro Arm, die zwischen den Armen an jeweils einer Geschlechtsöffnung enden. Seeigel haben zwischen drei und fünf Gonaden, die an den Interradien befestigt sind und nach außen münden. Bei den Seewalzen existiert nur eine Gonade, die sich im Körper verzweigt und in einem Geschlechtsgang mit Geschlechtsöffnung am Hinterende mündet. Da viele ausgestorbene Stachelhäuter ebenfalls nur eine Geschlechtsöffnung hatten, wird dieser Zustand als ursprünglich angesehen. ⓘ
Regeneration

Viele Stachelhäuter verfügen über große Regenerationsfähigkeiten. Bei vielen Arten kommt es zur Autotomie und zur Regeneration von Armen und Eingeweiden. Seegurken werfen oft Teile ihrer inneren Organe ab, wenn sie sich bedroht fühlen, und regenerieren sie im Laufe mehrerer Monate. Seeigel ersetzen ständig Stacheln, die sie durch Verletzungen verloren haben, während Seesterne und Seelilien ihre Arme leicht verlieren und regenerieren können. In den meisten Fällen kann ein einzelner abgetrennter Arm nicht zu einem neuen Seestern heranwachsen, wenn zumindest ein Teil der Scheibe fehlt. Bei einigen wenigen Arten kann jedoch ein einzelner Arm überleben und sich zu einem vollständigen Individuum entwickeln, und manchmal werden Arme absichtlich zum Zweck der ungeschlechtlichen Fortpflanzung abgetrennt. In Zeiten, in denen sie ihren Verdauungstrakt verloren haben, leben Seegurken von gespeicherten Nährstoffen und nehmen gelöste organische Stoffe direkt aus dem Wasser auf. ⓘ
Die Regeneration verlorener Teile erfolgt sowohl durch Epimorphose als auch durch Morphallaxe. Bei der Epimorphose bilden Stammzellen - entweder aus einem Reservepool oder durch Dedifferenzierung - ein Blastema und erzeugen neues Gewebe. Bei der morpholaktischen Regeneration werden vorhandene Gewebe bewegt und umgebaut, um verlorene Teile zu ersetzen. Auch die direkte Transdifferenzierung eines Gewebetyps in einen anderen wird beim Gewebeersatz beobachtet. ⓘ
Fortpflanzung
Sexuelle Fortpflanzung
Stachelhäuter werden je nach Art und Umweltbedingungen nach etwa zwei bis drei Jahren geschlechtsreif. Fast alle Arten haben getrennte männliche und weibliche Geschlechter, einige sind jedoch zwittrig. Die Eier und Spermien werden in der Regel ins offene Wasser entlassen, wo die Befruchtung stattfindet. Bei einigen Arten erfolgt die Freisetzung von Spermien und Eiern synchron, meist im Einklang mit dem Mondzyklus. Bei anderen Arten können sich die Tiere während der Fortpflanzungszeit zusammenfinden, was die Wahrscheinlichkeit einer erfolgreichen Befruchtung erhöht. Interne Befruchtung wurde bei drei Seesternarten, drei Schlangensternen und einer Tiefwasserseegurke beobachtet. Selbst in abgrundtiefen Gewässern, in die kein Licht eindringt, synchronisieren Stachelhäuter häufig ihre Fortpflanzungsaktivitäten. ⓘ
Einige Stachelhäuter brüten ihre Eier aus. Dies ist vor allem bei Kaltwasserarten üblich, bei denen die planktonischen Larven möglicherweise nicht genügend Nahrung finden. Diese zurückgehaltenen Eier sind in der Regel nicht sehr zahlreich und werden mit einem großen Eigelb versehen, um die sich entwickelnden Embryonen zu ernähren. Bei Seesternen kann das Weibchen die Eier in speziellen Beuteln, unter den Armen, unter ihrem gewölbten Körper oder sogar in ihrem Herzmagen tragen. Viele Schlangensterne sind Zwitter; sie brüten ihre Eier oft aus, meist in speziellen Kammern auf der Mundoberfläche, manchmal aber auch im Eierstock oder im Coelom. Bei diesen Seesternen und Schlangensternen erfolgt die Entwicklung in der Regel direkt bis zur erwachsenen Form, ohne dass ein zweiseitiges Larvenstadium durchlaufen wird. Einige Seeigel und eine Sanddollarart tragen ihre Eier in Höhlen oder in der Nähe ihres Afters und halten sie mit ihren Stacheln fest. Einige Seegurken benutzen ihre Wangententakel, um ihre Eier auf die Unterseite oder den Rücken zu übertragen, wo sie festgehalten werden. Bei einigen wenigen Arten werden die Eier im Coelom zurückbehalten, wo sie sich lebendig entwickeln und später durch Risse in der Körperwand austreten. Bei einigen Seelilien entwickeln sich die Embryonen in speziellen Brutbeuteln, in denen die Eier aufbewahrt werden, bis ein von einem Männchen freigesetztes Sperma sie findet. ⓘ
Ungeschlechtliche Fortpflanzung
- Siehe auch: Abschnitt "Regeneration" oben ⓘ

Eine Seesternart, Ophidiaster granifer, pflanzt sich ungeschlechtlich durch Parthenogenese fort. Bei einigen anderen Asterozoen pflanzen sich die erwachsenen Tiere ungeschlechtlich fort, bis sie geschlechtsreif sind und sich dann sexuell fortpflanzen. Bei den meisten dieser Arten erfolgt die ungeschlechtliche Fortpflanzung durch Querspaltung, bei der sich die Scheibe in zwei Teile teilt. Sowohl der verlorene Scheibenbereich als auch die fehlenden Arme wachsen nach, so dass ein Individuum Arme unterschiedlicher Länge haben kann. In der Zeit des Nachwachsens haben sie einige winzige Arme und einen großen Arm, weshalb sie oft als "Kometen" bezeichnet werden. ⓘ
Erwachsene Seegurken pflanzen sich ungeschlechtlich durch Querspaltung fort. Holothuria parvula wendet diese Methode häufig an, indem sie sich kurz vor der Mitte in zwei Teile spaltet. Die beiden Hälften regenerieren jeweils ihre fehlenden Organe über einen Zeitraum von mehreren Monaten, aber die fehlenden Genitalorgane entwickeln sich oft nur sehr langsam. ⓘ
Die Larven einiger Stachelhäuter sind zur ungeschlechtlichen Fortpflanzung fähig. Dies ist seit langem bei Seesternen und Schlangensternen bekannt, wurde aber in jüngerer Zeit auch bei einer Seegurke, einem Sanddollar und einem Seeigel beobachtet. Dies kann durch Autotomisierung von Teilen geschehen, die sich zu Sekundärlarven entwickeln, durch Knospung oder durch Querteilung. Autotomisierte Teile oder Knospen können sich direkt zu voll ausgebildeten Larven entwickeln, oder sie durchlaufen ein Gastrula- oder sogar ein Blastula-Stadium. Neue Larven können sich aus der präoralen Haube (einer hügelartigen Struktur über dem Mund), der seitlichen Körperwand, den postero-lateralen Armen oder ihren hinteren Enden entwickeln. ⓘ
Das Klonen ist für die Larve sowohl ressourcen- als auch entwicklungszeitaufwendig. Die Larven durchlaufen diesen Prozess, wenn reichlich Nahrung vorhanden ist oder die Temperaturbedingungen optimal sind. Das Klonen kann dazu dienen, die Gewebe zu nutzen, die normalerweise während der Metamorphose verloren gehen. Die Larven einiger Sanddollars klonen sich selbst, wenn sie gelösten Fischschleim entdecken, der die Anwesenheit von Fressfeinden anzeigt. Die ungeschlechtliche Vermehrung bringt viele kleinere Larven hervor, die besser vor planktivoren Fischen fliehen können, was darauf hindeutet, dass es sich bei diesem Mechanismus um eine Anpassung zum Schutz vor Fressfeinden handeln könnte. ⓘ
Entwicklung der Larven

Den Beginn der Entwicklung stellt eine Radiärfurchung dar mit annähernd äqualer Teilung. Es entwickelt sich erst eine Coeloblastula, aus der durch Einstülpung eine Gastrula wird. In den Hohlraum wandern Mesodermzellen ein, die später Skelettelemente bilden. Die sekundäre Leibeshöhle (Coelom) bildet sich durch Abschnürung von drei paarigen Blasen (Enterocoelie). ⓘ
Mit der Metamorphose bilden sich für die verschiedenen Taxa der Stachelhäuter unterschiedliche typische Larvenformen. Dabei handelt es sich bei den Pelmatozoa (Seelilien und Haarsterne) immer um die dotterreichen Doliolaria- oder Vitellaria-Larven, die nach ihrem Festsetzen ein fressfähiges Pentacrinusstadium bilden, aus der sich später das fertige Tier entwickelt. Die Grundlarvenform aller Eleutherozoa ist die Dipleurula-Larve, die sich bei den Seesternen über eine Bipinnaria-Larve und eine Brachiolaria-Larve zum adulten Tier entwickelt. Bei den Seegurken ist die erste Larvenform die Auricularia-Larve mit einem einheitlichen Wimpernband, aus der sich dann die Doliolaria mit drei bis fünf Wimpernringen entwickelt. Die auffälligsten Larven bilden die Seeigel und die Schlangensterne. Dabei handelt es sich um Pluteus-Larven mit langen Schwebestacheln. Die Larvenform wird dabei durch den Dottergehalt der Eizelle und die damit verbundene Lebensweise bestimmt. Larven, welche aus einer dotterarmen Eizelle hervorgehen leben in der Regel planktotroph, das heißt, sie ernähren sich im frei schwimmenden Larvenstadium von Organismen des Planktons. Im Gegensatz dazu leben Larven, welche aus einer dotterreichen Eizelle hervorgehen, lecithotroph, was bedeutet, dass sie sich vom eigenen Dotter ernähren und daher keinen Darm benötigen. Crinoida ausgenommen, da diese nur lecithotroph lebende Larvenformen aufweisen, weisen alle anderen Klassen sowohl Vertreter mit planktotroph- als auch Vertreter mit lecithotroph-lebenden Larven auf. ⓘ
In der Larvalentwicklung kommt es zur bereits angesprochenen Metamorphose, bei der aus den anfänglichen bilateralsymmetrischen Larven die fünfstrahligen Tiere entstehen. Den wesentlichen Teil dabei bildet das ursprünglich in drei paarigen Hohlräumen angelegte Coelom. Dieses entsteht durch eine beidseitige Abschnürung im Urdarmdach, also durch die bei den Deuterostomia übliche Enterocoelie, als Bläschen. Die Bläschen wachsen sich aus und teilen sich in Richtung Hinterpol in drei Abschnitte. Dabei bildet das Protocoel den obersten, das Mesocoel den mittleren und das Metacoel den hintersten Abschnitt. Das linke und das rechte Metacoel lagern sich als Somatocoel übereinander und bilden die eigentliche Leibeshöhle, die den Darm umgibt und die Geschlechtsdrüsen enthält. Aus dem linken Protocoel entwickelt sich das Axocoel, welches über einen Kanal und die Madreporenplatte nach außen mündet. Dabei bildet sich aus dem linken Mesocoel als Hydrocoel ein Ringkanal als Zentrum des Ambulacralsystems, der sich um den Darm legt und mit dem Axocoel über den Steinkanal verbunden ist. Aus dem Ringkanal wachsen Radiärkanäle in die späteren Radien ein. Das rechte Hydrocoel verschwindet vollständig und das rechte Axocoel wird auf eine kleine Dorsalblase reduziert. Bei den meisten Tieren bildet sich entsprechend die Oralseite auf der linken Körperseite. Abänderungen dieser Entwicklung gibt es bei den Seelilien, Haarsternen und den Seewalzen. ⓘ
Die Entwicklung beginnt mit einem zweiseitig symmetrischen Embryo, wobei sich zunächst eine Coeloblastula entwickelt. Mit der Gastrulation öffnet sich der "zweite Mund", durch den die Stachelhäuter zu den Deuterostomiern gehören, und das Mesoderm, das das Skelett beherbergen wird, wandert nach innen. Die sekundäre Körperhöhle, das Coelom, bildet sich durch die Aufteilung von drei Körperhöhlen. Die Larven sind oft planktonisch, aber bei einigen Arten bleiben die Eier im Inneren des Weibchens, während bei anderen das Weibchen die Larven ausbrütet. ⓘ
Verbreitung und Lebensraum

Stachelhäuter sind weltweit in allen Meeren und Ozeanen zu finden, einige Arten leben auch im Brackwasser. Dabei sind sie regional sehr häufig und stellen in vielen Bereichen die häufigsten Bewohner des Meeresbodens dar. Die verschiedenen Arten sind meistens spezifisch an bestimmte Umweltbedingungen angepasst und kommen entsprechend nur regional begrenzt vor. So gibt es unter den Stachelhäutern sehr viele Arten, die ausschließlich in den Tropen zu finden sind, andere leben in den polaren Gewässern der Arktis oder der Antarktis. Unter den tropischen Arten findet man viele, die immer an Korallenriffe gebunden sind, jedoch in allen Riffen im gesamten Indischen Ozean und Pazifischen Ozean vorkommen. ⓘ
Die meisten Stachelhäuter leben vollständig auf dem Meeresboden und auf anderen, ähnlichen Strukturen wie Felsstrukturen oder Korallenriffen. Viele Arten graben sich dabei auch in den Meeresboden ein. Nur wenige Seewalzenarten der Tiefsee sind freischwimmend und leben in den unteren Meeresschichten. Dabei sind alle Formen bis auf die sessilen Seelilien sehr mobil und können aktiv weite Strecken zurücklegen. Besonders die Seeigel und die Schlangensterne nutzen zur Verbreitung auch ihre freischwimmenden Larven, die eine weitere Streuung ermöglichen. Ihre Tiefenverbreitung ist wiederum artabhängig. Einige Schlangensterne leben zu tausenden in den Röhrensystemen von tropischen Schwämmen. Auch einige Seewalzen leben an anderen Organismen, etwa zwischen den Stacheln großer Seeigel in der Antarktis. Eine Seewalzenart befestigt ihren Körper sogar an der Haut von Tiefseefischen und lässt sich auf diese Weise transportieren. ⓘ
Die Arten der Flachwasserzonen und Meeresküsten kommen dabei selten in größeren Tiefen als 100 Meter vor. Tiefseearten finden sich in 1000 bis 5000 Metern Tiefe. Zu den Tiefseearten gehören auch die Seegänseblümchen, die ausschließlich in untergetauchtem Holz in der Tiefsee gefunden wurden. Aus Gebieten mit 10.000 Metern Tiefe und mehr sind bislang ausschließlich Seewalzenarten bekannt. Diese stellen hier in einigen Bereichen bis zu 90 Prozent aller lebenden Organismen dar. ⓘ
Stachelhäuter sind weltweit in fast allen Tiefen, Breitengraden und Umgebungen des Ozeans verbreitet. Ausgewachsene Tiere leben hauptsächlich benthisch auf dem Meeresboden, während Larven oft pelagisch als Plankton im offenen Ozean leben. Einige ausgewachsene Holothuroiden wie Pelagothuria sind jedoch pelagisch. Einige Seelilien sind pseudoplanktonisch, d. h. sie heften sich an schwimmende Baumstämme und Trümmer, obwohl dieses Verhalten im Paläozoikum am weitesten verbreitet war, bevor die Konkurrenz durch Organismen wie Seepocken das Ausmaß dieses Verhaltens einschränkte. ⓘ
Lebensweise
Fortbewegung

Stachelhäuter bewegen sich in erster Linie mit ihren Röhrenfüßen fort, manche Seeigel benutzen aber auch ihre Stacheln. Die Röhrenfüße haben in der Regel eine Spitze, die wie ein Saugnapf geformt ist, in dem durch Kontraktion der Muskeln ein Vakuum erzeugt werden kann. In Kombination mit einer gewissen Klebrigkeit, die durch die Absonderung von Schleim entsteht, sorgt dies für Haftung. Die Röhrenfüße kontrahieren und entspannen sich in Wellen, die sich entlang der haftenden Oberfläche bewegen, und das Tier bewegt sich langsam fort. ⓘ
Schlangensterne sind die beweglichsten unter den Stachelhäutern. Jeder der Arme kann die Symmetrieachse bilden, die entweder nach vorne oder nach hinten zeigt. Das Tier bewegt sich dann auf koordinierte Weise, angetrieben von den anderen vier Armen. Bei der Fortbewegung können die Antriebsarme entweder schlangenartige oder rudernde Bewegungen ausführen. Seesterne bewegen sich mit ihren Röhrenfüßen und halten ihre Arme fast still, auch bei Gattungen wie Pycnopodia, wo die Arme flexibel sind. Die Mundoberfläche ist mit Tausenden von Röhrenfüßen bedeckt, die sich zwar nicht im Takt, aber doch in einem metachronischen Rhythmus bewegen; in gewisser Weise sind die Röhrenfüße jedoch koordiniert, da das Tier gleichmäßig dahingleitet. Einige wühlende Seesterne haben an ihren Röhrenfüßen eher Spitzen als Saugnäpfe und können so schneller über den Meeresboden "gleiten". ⓘ
Seeigel benutzen ihre Röhrenfüße, um sich auf ähnliche Weise fortzubewegen wie Seesterne. Einige verwenden auch ihre gegliederten Stacheln, um sich vorwärts zu schieben oder zu hebeln oder um ihre Mundflächen vom Untergrund abzuheben. Wenn ein Seeigel umgekippt ist, kann er seine Röhrenfüße in einem Ambulakralbereich so weit ausfahren, dass sie in Reichweite des Substrats sind, und dann nacheinander Füße aus dem angrenzenden Bereich anbringen, bis er wieder aufgerichtet ist. Einige Arten bohren sich in Felsen, indem sie mit ihren Mundwerkzeugen an der Oberfläche schleifen. ⓘ
.jpg)
Seegurken sind im Allgemeinen träge Tiere. Viele können sich auf der Oberfläche des Meeresbodens bewegen oder sich durch peristaltische Bewegungen in Sand oder Schlamm eingraben; einige haben kurze Röhrenfüße an der Unterseite, mit denen sie wie ein Seestern kriechen können. Einige Arten ziehen sich mit ihren Wangententakeln fort, während andere durch peristaltische Bewegungen oder rhythmisches Biegen schwimmen können. Viele leben in Rissen, Höhlen und Höhlungen und bewegen sich kaum. Einige Tiefseearten sind pelagisch und können mit Schwimmhäuten, die Segel oder Flossen bilden, im Wasser schwimmen. ⓘ
Die meisten Seelilien sind beweglich, aber Seelilien sind sessil und mit Stielen an harten Substraten befestigt. Die Bewegung der meisten Seelilien beschränkt sich auf das Biegen der Stiele und das Ein- und Ausrollen der Arme; einige wenige Arten können sich auf dem Meeresboden kriechend fortbewegen. Die Seefedern sind ungebunden und leben meist in Spalten, unter Korallen oder in Schwämmen, wobei ihre Arme der einzige sichtbare Teil sind. Einige Seefedern tauchen nachts auf und setzen sich auf nahe gelegene Erhebungen, um die nahrungsbringenden Strömungen besser nutzen zu können. Viele Arten können über den Meeresboden "laufen", indem sie ihren Körper mit Hilfe ihrer Arme aufrichten, oder sie können mit Hilfe ihrer Arme schwimmen. Die meisten Arten von Seefedern sind jedoch weitgehend sesshaft und bewegen sich nur selten weit von ihrem gewählten Versteck weg. ⓘ
.jpg)

Seesterne und Seeigel bewegen sich vor allem durch ihre Ambulacralfüßchen sowie durch die Stacheln fort. Sie sind dadurch in der Lage, in jede Richtung zu wandern. Diese Füßchen werden auch als Saugnäpfchen eingesetzt, wodurch die Tiere auch an senkrechten Strukturen klettern können. Die Schlangensterne nutzen ihre Arme zur Fortbewegung, indem sie sie schlängelnd bewegen und in die Bewegungsrichtung schlagen. Auch die Seewalzen nutzen die Ambulacralfüßchen zur Fortbewegung. Dabei wandern sie meistens in Richtung des Vorderendes und unterstützen die Bewegung durch Kontraktionen ihres Körpers. Die Synaptidae verankern sich außerdem mit ihren klebrigen Tentakeln und ziehen sich dadurch vorwärts. ⓘ
Die Seelilien bewegen sich in der Regel gar nicht fort, obwohl es bei ihnen auch Arten gibt, die sich von ihrem Fixpunkt lösen und an anderer Stelle erneut festsetzen können. Die Haarsterne schwimmen, indem sie ihre Arme auf- und abwärts schlagen, doch auch sie sind meistens ortstreu. Eine schwimmende Fortbewegung kennt man außerdem bei einigen Schlangensternen und vor allem bei speziell abgeflachten Seewalzen der Tiefsee. Diese haben meist einen gelatinösen Körper und besitzen kein Skelett mehr. ⓘ
Sehr viele Arten der Stachelhäuter graben im Boden und bewegen sich entsprechend auch grabend fort. Dabei nutzen die Seeigel ihre Stacheln, Pedicellarien und Ambulacralfüßchen zum Graben, einige Arten haben außerdem die Möglichkeit, sich durch ihre Zähne und Stacheln in Felsgestein zu bohren. Die Seewalzen graben sich mit Hilfe ihrer Tentakel sowie kontrahierenden Bewegungen ein. Seeigel und Schlangensterne nutzen ihre Arme zum Graben. ⓘ
Die meisten Stachelhäuter haben eine Lebendposition, bei der die Mundöffnung zum Boden weist. Werden sie umgedreht, so versuchen sie durch Streckungen und Verdrehungen, die ursprüngliche Position wieder einzunehmen. ⓘ
Fütterung
Die Art der Ernährung ist bei den verschiedenen Stachelhäutern sehr unterschiedlich. Stachelhäuter und einige Schlangensterne sind in der Regel passive Filtrierer, die Schwebeteilchen aus dem vorbeifließenden Wasser aufsaugen. Die meisten Seeigel sind Weidegänger, Seegurken sind Ablagerungsfresser, und die meisten Seesterne sind aktive Jäger. ⓘ
Stachelhäuter fangen Nahrungspartikel mit den Röhrenfüßen an ihren ausgebreiteten Fiederblättern, transportieren sie in die Ambulakralrillen, wickeln sie in Schleim ein und befördern sie mit Hilfe der Flimmerhärchen, die die Rillen auskleiden, zum Mund. Die genauen Ernährungsbedürfnisse der Seelilien sind wenig erforscht, aber im Labor können sie mit Kieselalgen gefüttert werden. ⓘ
Korbsterne sind Suspensionsfresser, die ihre verzweigten Arme zum Aufsammeln von Zooplankton hochheben, während andere Schlangensterne mehrere Methoden der Nahrungsaufnahme anwenden. Einige sind Schwebfresser, die Nahrungspartikel mit Schleimfäden, Stacheln oder Röhrenfüßen an ihren erhobenen Armen festhalten. Andere sind Aasfresser und Detritusfresser. Wieder andere sind gefräßige Fleischfresser und können ihre Beute im Wasser mit einem plötzlichen Einkreisen durch ihre flexiblen Arme einfangen. Die Gliedmaßen biegen sich dann unter die Scheibe, um die Nahrung in die Kiefer und das Maul zu befördern. ⓘ
Viele Seeigel ernähren sich von Algen, wobei sie oft die dünne Algenschicht, die die Oberfläche von Felsen bedeckt, mit ihren spezialisierten Mundwerkzeugen, den so genannten Aristoteleslaternen, abkratzen. Andere Arten verschlingen kleinere Organismen, die sie mit ihren Röhrenfüßen fangen können. Sie können sich auch von toten Fischen und anderen tierischen Stoffen ernähren. Sanddollars können sich als Suspensionsfresser von Phytoplankton, Detritus, Algenstücken und der Bakterienschicht, die Sandkörner umgibt, ernähren. ⓘ
Seegurken sind oft mobile Ablagerungs- oder Suspensionsfresser, die mit ihren Mundwerkzeugen aktiv Nahrung aufnehmen und die Partikel dann einzeln in ihre Mundhöhle stopfen. Andere nehmen große Mengen an Sediment auf, absorbieren die organische Substanz und leiten die unverdaulichen mineralischen Partikel durch ihre Eingeweide weiter. Auf diese Weise stören und verarbeiten sie große Mengen an Substrat und hinterlassen oft charakteristische Sedimentkämme auf dem Meeresboden. Einige Seegurken leben infaunal in Höhlen, mit dem Vorderende nach unten und dem Anus an der Oberfläche, schlucken Sedimente und leiten sie durch ihren Darm. Andere Seegurken leben mit dem Vorderende nach oben und warten darauf, dass Detritus in die Eingänge der Höhlen fällt, oder sie harken mit ihren Mundwerkzeugen Trümmer von der Oberfläche in der Nähe ein. ⓘ
Fast alle Seesterne sind Detritus- oder Fleischfresser, aber einige wenige ernähren sich von Schwebstoffen. Kleine Fische, die an der Oberfläche landen, können von den Pedicilarien gefangen werden, und tote Tiere können erbeutet werden, aber die Hauptbeute sind lebende wirbellose Tiere, vor allem Muscheln und Weichtiere. Um sich von einer Muschel zu ernähren, bewegt sich der Seestern über sie, setzt seine Röhrenfüße an und übt Druck auf die Klappen aus, indem er seinen Rücken krümmt. Wenn sich ein kleiner Spalt zwischen den Ventilen gebildet hat, schiebt der Seestern einen Teil seines Magens in die Beute, scheidet Verdauungsenzyme aus und verflüssigt langsam die weichen Körperteile. Wenn sich der Adduktorenmuskel der Muschel entspannt, wird mehr Magen eingeführt, und wenn die Verdauung abgeschlossen ist, kehrt der Magen in seine übliche Position im Seestern zurück, mit der nun verflüssigten Muschelmahlzeit darin. Andere Seesterne stülpen den Magen aus, um sich von Schwämmen, Seeanemonen, Korallen, Detritus und Algenfilmen zu ernähren. ⓘ
Verteidigung gegen Raubtiere
.jpg)
Trotz ihres geringen Nährwerts und des hohen Anteils an unverdaulichem Kalzit werden Stachelhäuter von vielen Organismen gefressen, darunter Knochenfische, Haie, Eiderenten, Möwen, Krebse, Schnecken, andere Stachelhäuter, Seeotter, Polarfüchse und Menschen. Größere Seesterne ernähren sich von kleineren; die große Menge an Eiern und Larven, die sie produzieren, sind Teil des Zooplanktons, das von vielen Meeresbewohnern verzehrt wird. Seelilien hingegen sind relativ frei von Raubtieren. ⓘ
Zu den Abwehrmechanismen gegen Fressfeinde gehören Stacheln, Giftstoffe (inhärent oder durch die Röhrenfüße abgegeben) und das Abwerfen von klebrigen Fäden durch Seegurken. Obwohl die Stacheln der meisten Stachelhäuter stumpf sind, sind die des Dornenkronenseesterns lang und scharf und können eine schmerzhafte Stichwunde verursachen, da das Epithel, das sie bedeckt, ein Toxin enthält. Aufgrund ihres fängigen Bindegewebes, das sich schnell von einem schlaffen in einen starren Zustand verwandeln kann, sind Stachelhäuter sehr schwer aus Spalten zu entfernen. Einige Seegurken verfügen über ein Bündel von Cuvierien, die als lange, klebrige Fäden aus ihrem Anus ausgestoßen werden können, um einen Angreifer zu verwickeln und dauerhaft außer Gefecht zu setzen. Seegurken verteidigen sich gelegentlich, indem sie ihre Körperwand aufreißen und den Darm und die inneren Organe ausstoßen. Seesterne und Schlangensterne können bei einem Angriff eine Autotomie durchführen, bei der ein Arm abgetrennt wird; dies kann den Angreifer so lange ablenken, dass das Tier entkommen kann. Einige Seesternarten können bei Gefahr wegschwimmen. ⓘ
Den Hauptschutz der Stachelhäuter stellen die bereits erwähnten Stacheln der Seeigel sowie ihr hartes Skelett dar. Hinzu kommen bei einigen Seeigeln Gifte, die sie über ihre Pedicellarien abgeben können. Viele Seewalzenarten stoßen bei Gefahr ein Gewirr von klebrigen Fäden aus, die Cuvierschen Schläuche, andere produzieren mit den Holothurinen starke Gifte. ⓘ
Aggregation
Bei vielen Stachelhäutern kommt es zur Bildung von großen Ansammlungen, vor allem bei verschiedenen Seeigeln und Schlangensternen. Dieses Phänomen ist bereits fossil überliefert, da viele fossile Formen in großen Fossillagerstätten aus einer oder wenigen Arten gefunden werden. Diese Ansammlungen lassen sich vor allem auf verschiedene Umweltbedingungen zurückführen, die den Tieren optimale Lebensbedingungen bieten. Dabei spielt vor allem die Verteilung der Nahrung eine große Rolle, so finden sich große Ansammlungen von Schlangensternen häufig in Bereichen mit hohem Planktonvorkommen. ⓘ
Ökologie

Stachelhäuter sind zahlreiche wirbellose Tiere, deren erwachsene Tiere eine wichtige Rolle in benthischen Ökosystemen spielen, während die Larven ein wichtiger Bestandteil des Planktons sind. Zu den ökologischen Aufgaben der adulten Tiere gehören das Abgrasen von Seeigeln, die Verarbeitung von Sedimenten durch Herzseeigel und die Suspensions- und Ablagerungsnahrung von Seelilien und Seegurken. Einige Seeigel können sich in festes Gestein bohren, wodurch Felswände destabilisiert und Nährstoffe ins Meer abgegeben werden. Auch Korallenriffe werden auf diese Weise durchbohrt, aber die Akkumulationsrate von Karbonatmaterial ist oft größer als die vom Seeigel verursachte Erosion. Stachelhäuter binden etwa 0,1 Gigatonnen Kohlendioxid pro Jahr in Form von Kalziumkarbonat und leisten damit einen wichtigen Beitrag zum globalen Kohlenstoffkreislauf. ⓘ
Stachelhäuter haben manchmal große Populationsschwankungen, die Ökosysteme verändern können. So führte 1983 das Massensterben des tropischen Seeigels Diadema antillarum in der Karibik zu einem Wechsel von einem von Korallen dominierten Riffsystem zu einem von Algen dominierten. Seeigel gehören zu den wichtigsten Pflanzenfressern in Riffen, und in der Regel besteht ein feines Gleichgewicht zwischen den Seeigeln und dem Seetang und anderen Algen, von denen sie sich ernähren. Ein Rückgang der Zahl der Raubtiere (Otter, Hummer und Fische) kann zu einem Anstieg der Seeigelzahlen führen, was eine Überweidung der Seetangwälder zur Folge hat und zu einer algenarmen "Seeigelbarre" führt. Am Great Barrier Reef hat ein ungeklärter Anstieg der Zahl der Dornenkronenseesterne (Acanthaster planci), die lebendes Korallengewebe abweiden, die Korallensterblichkeit stark erhöht und die Artenvielfalt des Korallenriffs verringert. ⓘ
Nutzung durch den Menschen
Als Nahrung und Medizin

Die wirtschaftliche Bedeutung der Stachelhäuter ist vornehmlich lokal. So werden pro Jahr etwa 50.000 Tonnen Seeigel gefangen, von denen besonders in Japan, Peru und in Frankreich die Gonaden bzw. die Eier gegessen werden. In Japan sind die Eier als Uni bekannt und stellen eine Zutat der Sushi-Küche dar. Dabei wird der Geschmack als weich und schmelzend beschrieben. Die Qualität ist abhängig von der Farbe, die von hellgelb bis leuchtend orange reichen kann. ⓘ
Auch Seewalzen stellen in einigen Ländern Südostasiens eine Delikatesse dar. Am bekanntesten sind sie als Trepang oder bêche-de-mer in China und Indonesien. Die Seewalzen werden dafür von den (häufig giftigen) Eingeweiden befreit und getrocknet. Besonders beliebt sind die Ananas-Seewalze (Thelenota ananas) (susuhan) und die rote Halodeima edulis. Die starken Giftstoffe der Seewalzen, bekannt als Holothurine, sind leicht psychoaktiv, Nutzungen der entheogenen Wirkungen sind jedoch nicht bekannt. Allerdings konnte festgestellt werden, dass Holothurine hemmend auf die Wachstumsrate verschiedener Tumorzellen wirkt, wodurch es für die Krebsmedizin interessant wird. ⓘ
Seesterne gelangen gemahlen als Kalkzusatz in industrielles Tierfutter, jährlich werden für diesen Zweck etwa 4000 Tonnen der Tiere verwendet. Vor allem in Gebieten, in denen kommerziell Muscheln, vor allem Austern, gezüchtet werden, können Massenauftreten von Seesternen einen großen wirtschaftlichen Schaden anrichten. ⓘ

Im Jahr 2019 wurden 129.000 Tonnen Stachelhäuter geerntet. Die meisten davon waren Seegurken (158.000 Tonnen) und Seeigel (73.000 Tonnen). Diese werden hauptsächlich als Nahrungsmittel, aber auch in der traditionellen chinesischen Medizin verwendet. Seegurken gelten in einigen südostasiatischen Ländern als Delikatesse und laufen daher Gefahr, übermäßig geerntet zu werden. Beliebte Arten sind die Ananasrolle Thelenota ananas (susuhan) und die Rote Seegurke Holothuria edulis. Diese und andere Arten sind in China und Indonesien umgangssprachlich als bêche de mer oder trepang bekannt. Die Seegurken werden zwanzig Minuten lang gekocht und dann sowohl natürlich als auch später über einem Feuer getrocknet, wodurch sie einen rauchigen Geschmack erhalten. In China werden sie als Grundlage für gelatinöse Suppen und Eintöpfe verwendet. Vor allem in Japan und Frankreich werden sowohl die männlichen als auch die weiblichen Keimdrüsen der Seeigel verzehrt. Der Geschmack wird als weich und schmelzend beschrieben, wie eine Mischung aus Meeresfrüchten und Obst. Es wurden Versuche zur Zucht von Seeigeln unternommen, um einen Ausgleich für die Überfischung zu schaffen. ⓘ
In der Forschung
Aufgrund ihres robusten Larvenwachstums werden Seeigel häufig in der Forschung eingesetzt, insbesondere als Modellorganismen in der Entwicklungsbiologie und Ökotoxikologie. Strongylocentrotus purpuratus und Arbacia punctulata werden zu diesem Zweck in embryologischen Studien verwendet. Die große Größe und die Transparenz der Eier ermöglichen die Beobachtung der Spermien bei der Befruchtung der Eizellen. Das Armregenerationspotenzial von Schlangensternen wird im Zusammenhang mit dem Verständnis und der Behandlung neurodegenerativer Erkrankungen beim Menschen untersucht. ⓘ
Andere Verwendungen
Die kalkhaltigen Tests oder Schalen von Stachelhäutern werden von Landwirten in Gebieten, in denen kein Kalkstein zur Verfügung steht, als Kalkquelle genutzt, und einige werden bei der Herstellung von Fischmehl verwendet. Vier Tausend Tonnen der Tiere werden jährlich für diese Zwecke verwendet. Dieser Handel erfolgt oft in Zusammenarbeit mit Muschelzüchtern, für die die Seesterne eine große Bedrohung darstellen, weil sie ihre Zuchttiere fressen. Weitere Verwendungszwecke für die von ihnen gefangenen Seesterne sind die Herstellung von Tierfutter, die Kompostierung und die Herstellung von getrockneten Exemplaren für den Kunsthandwerkermarkt. ⓘ
Siehe auch
- Liste der prähistorischen Stachelhäuter-Gattungen - - ⓘ
Zitierte Texte
Ökologische Rolle und Gefahr

Stachelhäuter leben auf der einen Seite von organischem Bestandsabfall auf dem Meeresboden und von Pflanzenteilen, auf der anderen stellen sie effektive Räuber kleinerer Tiere dar. Damit sorgen sie für den Abbau von organischem Material und für die Regulation der Kleintierpopulationen. In Korallenriffen kontrollieren verschiedene Seesterne das Wachstum von Algenmatten und verhindern damit ein Zuwachsen der Korallenstöcke. Seewalzen sorgen für eine Veränderung des Meeresbodens ähnlich wie dies Regenwürmer auf dem Land tun. Sie fressen große Mengen Substrat und scheiden dieses nach der Darmpassage wieder aus. Dadurch wird das Substrat von organischen Stoffen befreit und zugleich chemisch und physikalisch verändert. Durch Grabtätigkeit durchmischen verschiedene grabende Seesterne, Sanddollars und Herz-Seeigel den Boden in Tiefen bis etwa 30 Zentimeter, Bohrseeigel bohren sogar Löcher in feste Gesteine. ⓘ
Die großen Mengen von Larven der Stachelhäuter stellen außerdem eine reichhaltige Futterquelle für andere Tierarten dar. Auch die ausgewachsenen Stachelhäuter werden von verschiedenen Tieren gefressen, etwa von Seeottern, Meeresvögeln und einigen Fischen. Einige Stachelhäuter, vor allem Seewalzen, stellen den Lebensraum für andere Tiere dar. So leben in den Körpern verschiedene Würmer, aber auch Krabben und Meeresschnecken. Besonders bekannt sind verschiedene Grundeln, die im Darm von Seewalzen leben und den After als Eingangsloch ihrer „Höhlen“ nutzen. Die meisten dieser Bewohner sind Kommensalen, einige leben jedoch auch parasitisch, so die Schnecken der Familie Eulimidae. ⓘ
Vor allem, wenn die Tiere überhandnehmen, kann ihr Fraßverhalten allerdings zu einem Ungleichgewicht führen. So führen beispielsweise die Massenbestände von Seesternen in einigen Tangwaldregionen zu einem Absterben der großen Tangpflanzen, andere Arten ernähren sich von Korallenpolypen und führen bei Massenauftreten zum Absterben von ganzen Riffen. ⓘ
Aufgrund ihres Kalkskelettes, welches empfindlich auf pH-Wert Veränderungen reagiert, kann die Entwicklung der Larvenform als Indikator bei der Erforschung der erhöhten Acidität der Meere aufgrund der globalen Erwärmung dienen. Experimente zeigten, dass eine Verringerung des pH-Wertes zu einer geringeren Larvengröße und zu einer Zunahme von Fehlbildungen während der Larvalentwicklung führte, während eine Zunahme der Temperatur zu einem stärkeren Wachstum der Larven führte und damit die geringere Larvengröße aufgrund des geringeren pH-Wertes teilweise kompensierte. Dies zeigt deutlich den intensiven Einfluss von Klimaveränderungen auf Echinodermaten. Es wurden auch Hinweise darauf beobachtet, dass die Verschmutzung der Meere durch Xenoöstrogen-haltige Abfälle Einfluss auf das Verhältnis der Geschlechterverteilung innerhalb einer Population einiger Stachelhäuter haben kann, was jedoch noch nicht eindeutig bewiesen werden konnte. ⓘ
Stachelhäuter und Menschen
Die meisten Menschen kennen die Stachelhäuter eher von der unangenehmen Seite: wenn man sich in Küstennähe – z. B. bei Felsen oder einem Riff – viele giftige Stacheln eines Seeigels eintritt. Wegen ihres feinen, ringförmigen Aufbaus brechen diese beim Entfernen meist ab – und es braucht dann Geduld (Herauseitern) oder den Arzt. Für die Küche einiger Länder sind Stachelhäuter jedoch besondere Delikatessen. Für Kinder sind ihre Skelette ein beliebtes Sammelobjekt. ⓘ