Blüte


Eine Blüte, manchmal auch als Blume oder Blüte bezeichnet, ist die reproduktive Struktur von Blütenpflanzen (Pflanzen der Abteilung Angiospermae). Die biologische Funktion einer Blüte besteht darin, die Fortpflanzung zu erleichtern, indem sie in der Regel einen Mechanismus für die Vereinigung von Spermien und Eiern bereitstellt. Blüten können die Auskreuzung (Verschmelzung von Spermien und Eiern verschiedener Individuen einer Population) infolge von Fremdbestäubung erleichtern oder die Selbstbestäubung (Verschmelzung von Spermien und Eiern derselben Blüte) ermöglichen, wenn es zu einer Selbstbestäubung kommt.
Es gibt zwei Arten der Bestäubung: Selbstbestäubung und Fremdbestäubung. Selbstbestäubung liegt vor, wenn der Pollen der Antheren auf der Narbe derselben Blüte oder einer anderen Blüte derselben Pflanze abgelagert wird. Fremdbestäubung liegt vor, wenn der Pollen von der Anthere einer Blüte auf die Narbe einer anderen Blüte einer anderen Art übertragen wird. Selbstbestäubung findet bei Blüten statt, bei denen Staubgefäß und Fruchtblatt gleichzeitig reifen und so angeordnet sind, dass der Pollen auf der Narbe der Blüte landen kann. Diese Bestäubung erfordert keine Investition der Pflanze, um Nektar und Pollen als Nahrung für die Bestäuber bereitzustellen. ⓘ
Einige Blüten produzieren Diasporen ohne Befruchtung (Parthenokarpie). Blüten enthalten Sporangien und sind der Ort, an dem sich Gametophyten entwickeln. Viele Blüten haben sich so entwickelt, dass sie für Tiere attraktiv sind, so dass sie als Vektoren für die Übertragung von Pollen dienen. Nach der Befruchtung entwickelt sich der Fruchtknoten der Blüte zu einer Frucht, die Samen enthält. ⓘ
Blumen dienen nicht nur der Fortpflanzung blühender Pflanzen, sondern werden von den Menschen seit langem bewundert und genutzt, um die Umwelt zu verschönern, aber auch als Objekte der Romantik, des Rituals, der Esoterik, der Hexerei, der Religion, der ganzheitlichen Medizin und als Nahrungsquelle. ⓘ

1. Blütenboden (Receptakulum)
2. Kelchblätter (Sepalen)
3. Kronblätter (Petalen)
4. Staubblätter (Stamina)
5. Stempel (Pistill) ⓘ
Die Blüte einer Pflanze ist ein nach Eintritt der Blühreife zu beobachtender unverzweigter Kurzspross mit begrenztem Wachstum, dessen Blätter indirekt oder direkt im Dienst der geschlechtlichen Fortpflanzung stehen: indirekt als Schutz- oder Anlockungsorgane (Blütenhülle), direkt durch die Bildung der Fortpflanzungsorgane (Staubblätter und Fruchtblätter). ⓘ

Etymologie
Der Begriff Blume stammt aus dem Mittelenglischen flour, das sich sowohl auf das gemahlene Korn als auch auf die reproduktive Struktur von Pflanzen bezog, bevor es sich im 17. Ursprünglich stammt es vom lateinischen Namen der italienischen Göttin der Blumen, Flora, ab. Das frühe Wort für Blume im Englischen war blossom, obwohl es sich heute nur auf die Blüten von Obstbäumen bezieht. ⓘ
Morphologie

Die Morphologie einer Blume, d. h. ihre Form und Struktur, kann in zwei Teile unterteilt werden: den vegetativen Teil, der aus nicht reproduktiven Strukturen wie den Blütenblättern besteht, und den reproduktiven oder sexuellen Teil. Eine stereotype Blüte besteht aus vier Arten von Strukturen, die an der Spitze eines kurzen Stiels oder einer Achse, dem sogenannten Rezeptor, befestigt sind. Jedes dieser Teile oder Blütenorgane ist in einer Spirale angeordnet, die als Wirtel bezeichnet wird. Die vier Hauptwirbel (von der Basis der Blüte oder dem untersten Knoten aufwärts) sind der Kelch, die Blumenkrone, das Androeceum und das Gynoeceum. Kelch und Krone bilden zusammen den nicht-reproduktiven Teil der Blüte, der Perianth genannt wird, und können in manchen Fällen nicht unterschieden werden. Ist dies der Fall, so werden sie als Tepalen bezeichnet. ⓘ
Die einzelnen Blütenorgane sind bei den Angiospermen innerhalb der Blüte in einer festgelegten Reihenfolge von unten nach oben, beziehungsweise, da die Blütenachse ja gestaucht ist, von außen nach innen angeordnet. In dieser Reihenfolge werden sie auch ontogenetisch (im Laufe der Blütenbildung) angelegt. ⓘ
Perianth
Kelch
Die Kelchblätter, die zusammen als Kelch bezeichnet werden, sind modifizierte Blätter, die am äußersten Wirtel der Blüte auftreten. Sie sind blattähnlich, d. h. sie haben eine breite Basis, Spaltöffnungen, Nebenblätter und Chlorophyll. Die Kelchblätter sind oft wachsartig und zäh und wachsen schnell, um die Blüte während ihrer Entwicklung zu schützen. Sie können laubabwerfend sein, wachsen aber meist weiter, um die Verbreitung der Früchte zu unterstützen. Wenn die Kelchblätter miteinander verwachsen sind, nennt man sie gamosepalig. ⓘ
Blumenkrone
Die Blütenblätter, die zusammen die Blumenkrone bilden, sind fast oder ganz faserlose blattähnliche Strukturen, die den innersten Wirtel des Perianths bilden. Sie sind oft zart und dünn und werden in der Regel gefärbt, geformt oder beduftet, um die Bestäubung zu fördern. Obwohl sie von der Form her den Blättern ähneln, sind sie eher mit den Staubblättern vergleichbar, da sie sich fast gleichzeitig bilden, ihr späteres Wachstum aber verzögert ist. Ist die Blumenkrone miteinander verwachsen, nennt man sie sympetal. ⓘ
Fortpflanzung

Androeceum


Mikrosporophylle sind sporangientragende Blätter. In den Sporangien (Pollensäcken) werden die männlichen Mikrosporen gebildet. Die Mikrosporophylle sind je nach Samenpflanzengruppe sehr unterschiedlich ausgebildet, und eine Homologie zu den Blättern (φύλλον phýllon „Blatt“) ist nicht gänzlich geklärt. Neutraler könnten sie daher auch Mikrosporangienträger genannt werden. Beim Ginkgo ist das Mikrosporophyll ein Stiel, an dessen Spitze zwei Pollensäcke hängen; die Mikrosporophylle stehen zu vielen schraubig an der Blütenachse. Bei den Koniferen ist die männliche Blüte zapfenartig und besteht aus vielen meist schraubig angeordneten Mikrosporophyllen; an jedem Mikrosporophyll stehen zwei bis 20 Pollensäcke. Bei den Palmfarnen (Cycadeen) sind die Mikrosporophylle schuppenförmig und tragen fünf bis 1000 Pollensäcke; die Mikrosporophylle stehen in der männlichen Blüte schraubig. Bei der Welwitschie tragen die sechs miteinander verwachsenen Mikrosporophylle je drei Pollensäcke. Bei Gnetum trägt das einzige Mikrosporophyll ein oder zwei endständige Pollensäcke. Bei Ephedra ist das Mikrosporophyll häufig gegabelt und trägt zwei bis acht Gruppen von meist zwei verwachsenen Pollensäcken. ⓘ
Die Mikrosporophylle der Angiospermen werden Staubblätter (Stamina) genannt. Ein Staubblatt ist meist in den Staubfaden (Filament) und den Staubbeutel (Anthere) gegliedert. Letzterer besteht aus zwei durch das Konnektiv verbundenen Theken zu je zwei Pollensäcken. Die Gesamtheit der Staubblätter wird Androeceum genannt. Die Anzahl der Staubblätter pro Blüte beträgt zwischen einem und rund 2000 Stück. ⓘ
Gynoeceum



Die Megasporophylle tragen die für Samenpflanzen charakteristischen Samenanlagen, in deren Innerem die eigentlichen weiblichen Fortpflanzungsorgane liegen, welche die pflanzlichen Eizellen enthalten. Auch die Megasporophylle könnten neutraler als Megasporangienträger bezeichnet werden. Nur bei den Angiospermen heißen die Megasporophylle Fruchtblätter. ⓘ
Bei den Palmfarnen stehen die Samenanlagen eindeutig auf Blättern, sie sind phyllospor. Das schuppen- oder schildförmige Megasporophyll ist deutlich gestielt und trägt am unteren Rand der Spreite zwei Samenanlagen, bei Cycas sind es bis zu acht Samenanlagen entlang der Rhachis. Bei den Koniferen befinden sich ein bis 20 Samenanlagen auf der Oberfläche einer flächig ausgebildeten Samenschuppe. Die Samenschuppe ist ein modifizierter Kurztrieb. Selten stehen die Samenanlagen bei Koniferen endständig an Kurztrieben, wie bei der Eibe (Taxus). Ginkgo bildet an der Spitze eines gegabelten Stieles zwei Samenanlagen. Die Gnetopsida bilden pro Blüte nur eine endständige Samenanlage. ⓘ
Bei den Angiospermen sind die Samenanlagen in das Fruchtblatt (Karpell) eingeschlossen, daher auch der Name „Bedecktsamer“. Die Gesamtheit der Fruchtblätter bildet das Gynoeceum. Die Anzahl der Fruchtblätter pro Blüte liegt zwischen einem und rund 2000. Meist gliedert sich ein Fruchtblatt in eine Stielzone und eine Schlauchzone, der hohle Bereich, in dem die Samenanlagen liegen (auch Ovar genannt). Die Samenanlagen setzen an Plazenten an. Darüber schließt sich häufig ein Griffel an, an dem sich die Narbe befindet, die Empfängnisfläche für den Pollen. Die Narbe ist durch einen Transmissionskanal oder -gewebe mit den Samenanlagen verbunden. Die Fruchtblätter können frei sein (apokarpes oder chorikarpes Gynoeceum) oder miteinander verwachsen sein (coenokarp). Bei einem verwachsenen Gynoeceum spricht man von einem Stempel (Pistill). Sterile Blüten besitzen höchstens reduzierte Staub- und Fruchtblätter und haben häufig die Funktion eines Schauapparates. ⓘ
Variation
Obwohl diese Anordnung als "typisch" gilt, gibt es bei den Pflanzenarten eine große Vielfalt an Blütenstrukturen. Die vier Hauptteile einer Blüte werden im Allgemeinen durch ihre Position am Blütenboden und nicht durch ihre Funktion definiert. Vielen Blüten fehlen einige Teile, oder Teile können in andere Funktionen umgewandelt werden oder wie ein typischer Teil aussehen. In einigen Familien, wie den Hahnenfußgewächsen, sind die Blütenblätter stark reduziert, und bei vielen Arten sind die Kelchblätter bunt und blütenblattartig. Andere Blumen haben modifizierte Staubgefäße, die blütenblattähnlich sind; die gefüllten Blüten von Pfingstrosen und Rosen bestehen meist aus blütenblattähnlichen Staubgefäßen. ⓘ
Viele Blüten weisen eine Symmetrie auf. Wenn das Perianth von einem beliebigen Punkt aus durch die Mittelachse halbiert wird und symmetrische Hälften entstehen, wird die Blüte als aktinomorph oder regelmäßig bezeichnet. Dies ist ein Beispiel für radiale Symmetrie. Wenn die Blüten halbiert werden und nur eine Linie entsteht, die symmetrische Hälften hervorbringt, wird die Blüte als unregelmäßig oder zygomorph bezeichnet. Wenn sie in seltenen Fällen überhaupt keine Symmetrie aufweisen, werden sie asymmetrisch genannt. ⓘ
Die Blüten können an ihrer Basis direkt mit der Pflanze verbunden sein (sessil - der tragende Stängel oder Stiel ist stark reduziert oder fehlt). Der Stiel oder Stängel, der eine Blüte oder einen Blütenstand trägt, wird als Blütenstiel bezeichnet. Wenn ein Blütenstiel mehr als eine Blüte trägt, werden die Stiele, die jede Blüte mit der Hauptachse verbinden, als Blütenstiele bezeichnet. Die Spitze eines Blütenstiels bildet eine endständige Verdickung, die als Torus oder Receptaculum bezeichnet wird. ⓘ
Bei den meisten Arten haben die einzelnen Blüten sowohl Stempel als auch Staubgefäße. Diese Blüten werden von Botanikern als perfekt, bisexuell oder zwittrig beschrieben. Bei einigen Pflanzenarten sind die Blüten jedoch unvollkommen oder eingeschlechtlich: Sie haben entweder nur männliche (Staubgefäße) oder weibliche (Stempel) Teile. Im letzteren Fall, wenn eine einzelne Pflanze entweder weiblich oder männlich ist, wird die Art als zweihäusig betrachtet. Wenn jedoch eingeschlechtliche männliche und weibliche Blüten auf derselben Pflanze erscheinen, wird die Art als einhäusig bezeichnet. Viele Blüten haben Nektarien, das sind Drüsen, die eine zuckerhaltige Flüssigkeit produzieren, um Bestäuber anzulocken. Sie werden nicht als eigenständiges Organ betrachtet. ⓘ
Blütenstand

Bei den Arten, die mehr als eine Blüte an einer Achse haben, nennt man die Gesamtheit der Blüten einen Blütenstand. Manche Blütenstände bestehen aus vielen kleinen Blüten, die in einer Formation angeordnet sind, die einer einzigen Blüte ähnelt. Ein typisches Beispiel hierfür sind die meisten Vertreter der sehr großen Korbblütler (Asteraceae). Ein einzelnes Gänseblümchen oder eine Sonnenblume beispielsweise ist keine Blüte, sondern ein Blütenstand, der aus zahlreichen Blüten (oder Blütchen) besteht. Ein Blütenstand kann spezialisierte Stängel und modifizierte Blätter enthalten, die als Hochblätter bezeichnet werden. ⓘ
Blütendiagramme und Formeln
Eine Blütenformel ist eine Möglichkeit, die Struktur einer Blüte mit bestimmten Buchstaben, Zahlen und Symbolen darzustellen und so wesentliche Informationen über die Blüte in einer kompakten Form zu präsentieren. Sie kann ein Taxon darstellen, wobei in der Regel die Anzahl der verschiedenen Organe oder eine bestimmte Art angegeben wird. Blütenformeln wurden im frühen 19. Jahrhundert entwickelt und ihre Verwendung ist seitdem zurückgegangen. Prenner et al. (2010) haben eine Erweiterung des bestehenden Modells entwickelt, um die Beschreibungsmöglichkeiten der Formel zu erweitern. Das Format der Blütenformeln ist in den verschiedenen Teilen der Welt unterschiedlich, aber sie vermitteln dieselben Informationen. ⓘ
Die Struktur einer Blume kann auch mit Hilfe von Blumendiagrammen ausgedrückt werden. Die Verwendung schematischer Diagramme kann lange Beschreibungen oder komplizierte Zeichnungen ersetzen, um sowohl die Struktur als auch die Entwicklung der Blume zu verstehen. Solche Diagramme können wichtige Merkmale von Blumen zeigen, einschließlich der relativen Positionen der verschiedenen Organe, einschließlich des Vorhandenseins von Verschmelzung und Symmetrie, sowie strukturelle Details. ⓘ
Blütenachse
An der Blütenachse (Blütenboden oder Receptaculum) setzen die Blütenblätter an. Sie kann gestaucht, gestreckt und unterschiedlich geformt sein. Es können einzelne oder mehrere Wirtel durch interkalare Meristeme emporgehoben werden: Beispielsweise beim Kapernstrauch wird das Gynoeceum emporgehoben (Gynophor), bei den Passionsblumen stehen Gynoeceum und Androeceum erhaben (Androgynophor), und bei vielen Nelkengewächsen sind die Krone, das Androeceum das Gynoeceum emporgehoben (Anthophor). ⓘ
Es können auch ringförmige Bereiche um das Zentrum herum emporgehoben werden. Ein Beispiel ist ein Stamen-Corollentubus, wie er bei vielen Korbblütlern typisch ist, oder der Kelch-Kronbecher vieler Kürbisgewächse. Eine verbreiterte Blütenachse heißt Blütenbecher (Hypanthium). ⓘ
Werden alle Teile außer dem Fruchtknoten emporgehoben, entsteht eine Blütenröhre, welche die Blütenhülle und die Staubblätter trägt. Es entsteht ein perigyner Blütenbecher. Der Fruchtknoten wird dann als mittelständig bezeichnet. Ein oberständiger Fruchtknoten steht frei auf der Blütenachse. Wenn bei einem Blütenbecher die Außenseiten der Fruchtblätter miteinbezogen werden, entsteht ein unterständiger Fruchtknoten, das bedeutet er steht unterhalb der Blütenhülle so beispielsweise bei den Rosen. ⓘ
Blütenhülle
Die Blütenhülle (Perianth) sind sterile Blätter, die jedoch eindeutig zur Blüte gehören. Eine Blütenhülle wird nur von Gnetopsida und Angiospermen gebildet. Sind alle Blütenhüllblätter einheitlich gestaltet (homoiochlamydeisch), wird die Blütenhülle als Perigon oder einfaches Perianth bezeichnet, die Blätter als Tepalen (etwa bei der Tulpe). Dabei kann das Perigon aus einem Kreis (haplo- oder monochlamydeisch), zwei oder mehr Kreisen oder mehreren Schraubenumläufen bestehen; mehrfaches Perigon. Ein doppeltes Perianth (di-, heterochlamydeische Blüten) besteht aus ungleichartigen Blütenhüllblättern. Die äußeren, meist grünen sind die Kelchblätter (Sepalen) und bilden den Kelch (Calyx), die inneren, häufig auffallend gefärbten Blätter sind die Kronblätter (Petalen) und bilden die Krone (Corolla). Die evolutive Herkunft der Blütenhülle wird je nach Verwandtschaftsgruppe aus Hochblättern oder aus Staubblättern gedeutet. ⓘ
Nektarien
Nektarien sind Drüsen, die Nektar als Lockspeise für die Blütenbestäuber bilden. Dieser besteht meist aus einer Zuckerlösung, die auch Proteine, Aminosäuren und Vitamine enthalten kann. Seltener werden Öle gebildet (etwa beim Gilbweiderich, Lysimachia). Nektarien können von der Blütenhülle, den Staubblättern, dem Fruchtknoten, der Blütenachse, aber auch außerhalb der eigentlichen Blüte (extrafloral) gebildet werden. ⓘ
Entwicklung
Eine Blüte entwickelt sich an einem modifizierten Spross oder einer Achse aus einem determinierten apikalen Meristem (determiniert bedeutet, dass die Achse bis zu einer bestimmten Größe wächst). Sie hat komprimierte Internodien, die Strukturen tragen, die in der klassischen Pflanzenmorphologie als stark modifizierte Blätter interpretiert werden. Detaillierte Entwicklungsstudien haben jedoch gezeigt, dass die Staubgefäße oft mehr oder weniger wie modifizierte Stängel (Caulome) entstehen, die in einigen Fällen sogar Verzweigungen ähneln können. Berücksichtigt man die gesamte Vielfalt in der Entwicklung des Androeceums von Blütenpflanzen, so ergibt sich ein Kontinuum zwischen modifizierten Blättern (Phyllomen), modifizierten Stängeln (Caulomen) und modifizierten Verzweigungen (Sprossen). ⓘ
Übergang
Die Blüteninduktion ist die Umsteuerung der Pflanze bzw. des Meristems vom vegetativen Wachstum zur Blütenbildung. Auslöser können innere (endogene) oder äußere (exogene) Faktoren sein. Die Induktion ist irreversibel, kann also nicht rückgängig gemacht oder gestoppt werden. Es gibt vier wichtige Signalwege:
- Endogene oder autonome Auslöser sind vorhanden, aber ihrer Natur nach nicht näher bekannt. Bei vielen Arten ist jedoch ein bestimmtes Alter oder eine bestimmte Größe nötig, bevor sie blühen, etwa bei vielen Bäumen. Bei der Erbse ist der Blühzeitpunkt je nach Sorte genetisch fixiert. Spätblühende Ökotypen der Acker-Schmalwand (Arabidopsis thaliana) bilden ab einem bestimmten Alter autonom Blüten, wenn zuvor keine Induktion durch Umweltfaktoren erfolgte. Die Umschaltung auf Blütenbildung erfolgt durch eine Aktivierung von Heterochronie-Genen wie FCA (Flowering Control Arabidopsis).
- Bei den meisten Pflanzen wird der Blühzeitpunkt durch äußere Faktoren festgelegt. Die wichtigsten Faktoren sind dabei die Tageslänge und Kälteperioden (Vernalisation). Bei der Tageslänge werden Kurztagspflanzen und Langtagpflanzen unterschieden, die für die Blühinduktion bestimmte Tageslängen benötigen. Eine Rolle als Rezeptor spielt dabei das Phytochrom-System der Blätter, ebenso das Cryptochrom. Auch das Protein FKF1 dient als Photorezeptor und misst die Tageslänge. Diese Systeme beeinflussen das Constans-Gen (CO), dessen Exprimierung alleine schon ausreicht, die Blüteninduktion auszulösen. Als Signal von den Blättern an die Blüten wurde lange ein Blütenhormon Florigen postuliert, dessen Identität jedoch lange unbekannt blieb. Das Gen FT (siehe unten) ist ein Kandidat dafür, da es zwar in den Blättern transkribiert wird, aber in den Blütenmeristemen wirkt.
- Besonders monokarpe Pflanzen (die nur einmal blühen) benötigen eine Vernalisation, eine längere Kälteperiode, um blühen zu können. Bekannte Gene, die bei Kälte aktiviert werden, sind Frigida (FRI) bei Arabidopsis und VRN1 und VRN2 im Winterweizen. Deren Genprodukte hemmen das Gen FLC, das seinerseits die Blütenbildung unterdrückt. In Summe wird also durch FRI oder VRN die Blütenbildung ermöglicht, wenn auch nicht ausgelöst. Es sind meist noch weitere Faktoren zur Auslösung nötig.
- Bestimmte Pflanzenhormone, wie etwa die Gibberelline, sind wichtig in der Blüh-Stimulation in der Abwesenheit von Langtagbedingungen, der Wirkmechanismus ist jedoch noch unbekannt. ⓘ
Der Übergang zur Blüte ist eine der wichtigsten Phasenänderungen, die eine Pflanze während ihres Lebenszyklus durchläuft. Der Übergang muss zu einem Zeitpunkt erfolgen, der für die Befruchtung und die Bildung von Samen günstig ist, um einen maximalen Reproduktionserfolg zu gewährleisten. Um diesen Anforderungen gerecht zu werden, ist eine Pflanze in der Lage, wichtige endogene und umweltbedingte Signale zu interpretieren, wie z. B. Veränderungen im Gehalt an Pflanzenhormonen und jahreszeitlich bedingte Temperatur- und Photoperiodenänderungen. Viele mehrjährige und die meisten zweijährigen Pflanzen benötigen eine Vernalisation, um zu blühen. Die molekulare Interpretation dieser Signale erfolgt durch die Übertragung eines komplexen Signals, das als Florigen bekannt ist und an dem eine Reihe von Genen beteiligt sind, darunter Constans, Flowering Locus C und Flowering Locus T. Florigen wird in den Blättern unter fortpflanzungsfreundlichen Bedingungen produziert und wirkt in Knospen und wachsenden Spitzen, um eine Reihe verschiedener physiologischer und morphologischer Veränderungen hervorzurufen.

Der erste Schritt des Übergangs ist die Umwandlung der vegetativen Stängelprimordien in Blütenprimordien. Dies geschieht durch biochemische Veränderungen, die die zelluläre Differenzierung von Blatt-, Knospen- und Stängelgewebe in Gewebe verändern, das zu den Fortpflanzungsorganen heranwächst. Das Wachstum des mittleren Teils der Stängelspitze hört auf oder flacht ab, und an den Seiten entwickeln sich Ausstülpungen in Form von Quirlen oder Spiralen um die Außenseite des Stängelendes. Aus diesen Ausstülpungen entwickeln sich die Kelchblätter, Blütenblätter, Staubgefäße und Fruchtblätter. Wenn dieser Prozess einmal begonnen hat, kann er bei den meisten Pflanzen nicht mehr rückgängig gemacht werden, und die Stängel bilden Blüten aus, selbst wenn der Beginn der Blütenbildung von bestimmten Umweltfaktoren abhängig war. ⓘ
Entwicklung der Organe
Das ABC-Modell ist ein einfaches Modell, das die Gene beschreibt, die für die Entwicklung der Blüten verantwortlich sind. Drei Genaktivitäten interagieren auf kombinatorische Weise, um die Entwicklungsidentitäten der primordialen Organe innerhalb des Blütenapikalmeristems zu bestimmen. Diese Genfunktionen werden als A, B und C bezeichnet. A-Gene werden nur im äußeren und untersten Abschnitt des Apikalmeristems exprimiert, der sich zu einem Wirtel von Kelchblättern entwickelt. Im zweiten Wirtel werden sowohl A- als auch B-Gene exprimiert, was zur Bildung von Blütenblättern führt. Im dritten Wirtel wirken B- und C-Gene zusammen, um Staubblätter zu bilden, und in der Mitte der Blüte führen nur C-Gene zur Bildung von Fruchtblättern. Das Modell basiert auf Studien über abnorme Blüten und Mutationen bei Arabidopsis thaliana und dem Löwenmäulchen (Antirrhinum majus). Wenn beispielsweise die Funktion des Gens B verloren geht, bilden sich bei den mutierten Blüten im ersten Wirtel wie üblich Kelchblätter, aber auch im zweiten Wirtel anstelle der normalen Blütenblattbildung. Im dritten Wirtel wird durch das Fehlen der B-Funktion, aber das Vorhandensein der C-Funktion der vierte Wirtel nachgeahmt, was zur Bildung von Fruchtblättern auch im dritten Wirtel führt. ⓘ
Funktion
Der Hauptzweck einer Blüte ist die Fortpflanzung des Individuums und der Art. Alle Blütenpflanzen sind heterosporös, das heißt, jede einzelne Pflanze produziert zwei Arten von Sporen. Mikrosporen werden durch Meiose in den Staubbeuteln und Megasporen in den Eizellen im Fruchtknoten gebildet. Die Staubbeutel bestehen in der Regel aus vier Mikrosporangien, und die Samenanlage ist ein umhülltes Megasporangium. Beide Arten von Sporen entwickeln sich innerhalb der Sporangien zu Gametophyten. Wie bei allen heterosporösen Pflanzen entwickeln sich auch die Gametophyten innerhalb der Sporen, d. h. sie sind endosporisch. ⓘ
Bei den meisten Pflanzenarten haben die einzelnen Blüten sowohl funktionsfähige Fruchtblätter als auch Staubgefäße. Die Botaniker bezeichnen diese Blüten als perfekt oder bisexuell und die Arten als zwittrig. Bei einer Minderheit von Pflanzenarten fehlt den Blüten das eine oder das andere Fortpflanzungsorgan, und sie werden als unvollkommen oder eingeschlechtlich bezeichnet. Wenn die einzelnen Pflanzen einer Art jeweils eingeschlechtliche Blüten beider Geschlechter haben, ist die Art einhäusig. Haben die einzelnen Pflanzen jedoch nur eingeschlechtliche Blüten desselben Geschlechts, so ist die Art zweihäusig. ⓘ
Bestäubung


Der Hauptzweck der Blüte ist die Fortpflanzung. Da die Blüten die Fortpflanzungsorgane der Pflanze sind, vermitteln sie die Verbindung zwischen den im Pollen enthaltenen Spermien und den im Fruchtknoten enthaltenen Eizellen. Die Bestäubung ist die Bewegung des Pollens von den Staubbeuteln zur Narbe. Normalerweise wird der Pollen von einer Pflanze auf eine andere übertragen, was als Fremdbestäubung bezeichnet wird, aber viele Pflanzen sind auch in der Lage, sich selbst zu bestäuben. Die Fremdbestäubung wird bevorzugt, weil sie eine genetische Variation ermöglicht, die zum Überleben der Art beiträgt. Viele Blumen sind daher für die Bestäubung auf äußere Faktoren angewiesen, wie Wind, Wasser, Tiere und insbesondere Insekten. Aber auch größere Tiere wie Vögel, Fledermäuse und sogar einige Zwergsäugetiere können zum Einsatz kommen. Um dies zu erreichen, sind die Blumen so gestaltet, dass sie die Übertragung von Pollen von einer Pflanze auf eine andere der gleichen Art fördern. Der Zeitraum, in dem dieser Prozess stattfinden kann (wenn die Blüte voll entfaltet und funktionsfähig ist), wird als Anthese bezeichnet, weshalb die Erforschung der Bestäubungsbiologie als Anthekologie bezeichnet wird. ⓘ
Blühende Pflanzen stehen in der Regel unter dem evolutionären Druck, die Übertragung ihrer Pollen zu optimieren, was sich in der Regel in der Morphologie der Blüten und im Verhalten der Pflanzen widerspiegelt. Die Übertragung von Pollen zwischen Pflanzen kann über eine Reihe von "Vektoren" oder Methoden erfolgen. Etwa 80 % der Blütenpflanzen nutzen biotische, also lebende Vektoren. Andere verwenden abiotische oder nicht lebende Vektoren, und einige Pflanzen nutzen mehrere Vektoren, aber die meisten sind hoch spezialisiert. ⓘ
Obwohl einige Pflanzen zwischen diesen Gruppen liegen oder auch außerhalb, lassen sich die meisten Blüten in die folgenden zwei großen Gruppen von Bestäubungsmethoden einteilen: ⓘ
Biotische Bestäubung
Blumen, die biotische Vektoren verwenden, locken Insekten, Fledermäuse, Vögel oder andere Tiere an und nutzen sie, um den Pollen von einer Blüte zur nächsten zu übertragen. Oft sind sie in ihrer Form spezialisiert und haben eine Anordnung der Staubgefäße, die sicherstellt, dass die Pollenkörner auf den Körper des Bestäubers übertragen werden, wenn dieser auf der Suche nach seinem Lockstoff (wie Nektar, Pollen oder einem Partner) landet. Auf der Suche nach dem Lockstoff von vielen Blüten derselben Art überträgt der Bestäuber den Pollen auf die Narben aller Blüten, die er besucht, und zwar mit der gleichen Präzision. Viele Blüten beruhen auf der einfachen Nähe der Blütenteile zueinander, um die Bestäubung zu gewährleisten, während andere Blüten aufwendig gestaltet sind, um die Bestäubung sicherzustellen und eine Selbstbestäubung zu verhindern. Blumen nutzen Tiere wie Insekten (Entomophilie), Vögel (Ornithophilie), Fledermäuse (Chiropterophilie), Eidechsen und sogar Schnecken und Nacktschnecken (Malakophilie). ⓘ
Methoden der Anziehung

Da Pflanzen nicht von einem Standort zum anderen wandern können, haben sich viele Blumen so entwickelt, dass sie Tiere anlocken, die den Pollen zwischen Individuen in verstreuten Populationen übertragen. Am häufigsten werden Blumen von Insekten bestäubt, die als entomophil bezeichnet werden, was im Griechischen wörtlich "insektenliebend" bedeutet. Um diese Insekten anzulocken, haben Blumen an verschiedenen Stellen Drüsen, die Nektarien genannt werden und Tiere auf der Suche nach nahrhaftem Nektar anlocken. Vögel und Bienen haben ein Farbsehvermögen, das es ihnen ermöglicht, "bunte" Blumen aufzusuchen. Einige Blüten haben Muster, die so genannten Nektarleitlinien, die den Bestäubern zeigen, wo sie nach Nektar suchen müssen. Sie sind möglicherweise nur unter ultraviolettem Licht sichtbar, das für Bienen und einige andere Insekten sichtbar ist. ⓘ
Blumen locken Bestäuber auch durch ihren Duft an, obwohl nicht alle Blumendüfte für den Menschen ansprechend sind. Einige Blumen werden von Insekten bestäubt, die von verrottetem Fleisch angezogen werden, und haben Blüten, die nach toten Tieren riechen. Diese Blumen werden oft als Aasblumen bezeichnet, darunter Pflanzen der Gattung Rafflesia und der Aronstab. Blumen, die von nächtlichen Besuchern wie Fledermäusen und Motten bestäubt werden, konzentrieren sich wahrscheinlich auf den Duft, um Bestäuber anzulocken, weshalb die meisten dieser Blumen weiß sind. ⓘ
Die Blüten sind auch in ihrer Form spezialisiert und haben eine Anordnung der Staubgefäße, die sicherstellt, dass die Pollenkörner auf die Körper der Bestäuber übertragen werden, wenn diese auf der Suche nach ihrem Lockstoff landen. Andere Blumen nutzen Mimikry oder Pseudokopulation, um Bestäuber anzulocken. Viele Orchideen zum Beispiel bilden Blüten aus, die in Farbe, Form und Duft weiblichen Bienen oder Wespen ähneln. Die Männchen wandern auf der Suche nach einer Partnerin von einer Blüte zur nächsten und bestäuben die Blüten. ⓘ
Beziehungen zwischen Bestäubern
Viele Blumen stehen in enger Beziehung zu einem oder wenigen spezifischen Bestäuberorganismen. Viele Blumen ziehen zum Beispiel nur eine bestimmte Insektenart an und sind daher für eine erfolgreiche Fortpflanzung auf dieses Insekt angewiesen. Diese enge Beziehung ist ein Beispiel für Koevolution, da sich Blume und Bestäuber über einen langen Zeitraum hinweg gemeinsam entwickelt haben, um die Bedürfnisse des jeweils anderen zu erfüllen. Diese enge Beziehung verstärkt jedoch die negativen Auswirkungen des Aussterbens, da das Aussterben eines der beiden Mitglieder einer solchen Beziehung mit ziemlicher Sicherheit auch das Aussterben des anderen Mitglieds bedeuten würde. ⓘ
Abiotische Bestäubung


Blumen, die abiotische oder nicht lebende Vektoren verwenden, nutzen den Wind oder, viel seltener, Wasser, um Pollen von einer Blüte zur nächsten zu transportieren. Bei windverbreiteten (anemophilen) Arten werden die winzigen Pollenkörner durch den Wind zu anderen Blüten getragen, manchmal über viele tausend Kilometer. Häufige Beispiele sind Gräser, Birken und viele andere Arten aus der Ordnung der Fagales, Ambrosia und viele Seggen. Sie müssen keine Bestäuber anlocken und neigen daher nicht dazu, große, auffällige oder farbenfrohe Blüten zu bilden, und haben weder Nektarien noch einen auffälligen Duft. Aus diesem Grund haben die Pflanzen in der Regel viele tausend winzige Blüten mit vergleichsweise großen, gefiederten Narben, um die Wahrscheinlichkeit zu erhöhen, dass Pollen empfangen wird. Während der Pollen entomophiler Blüten in der Regel groß, klebrig und reich an Proteinen ist (als "Belohnung" für Bestäuber), ist der Pollen anemophiler Blüten in der Regel kleinkörnig, sehr leicht, glatt und von geringem Nährwert für Insekten. Damit der Wind den Pollen effektiv aufnehmen und transportieren kann, sind die Antheren der Blüten in der Regel lose am Ende langer dünner Filamente befestigt, oder der Pollen bildet sich um ein Kätzchen, das sich im Wind bewegt. Seltenere Formen sind einzelne Blüten, die vom Wind bewegt werden können (hängend), oder, noch seltener, die Antheren explodieren und geben den Pollen in den Wind ab. ⓘ
Die Bestäubung durch Wasser (Hydrophilie) ist eine viel seltenere Methode, die nur bei etwa 2 % der abiotisch bestäubten Blüten vorkommt. Häufige Beispiele hierfür sind Calitriche autumnalis, Vallisneria spiralis und einige Seegräser. Ein Merkmal, das die meisten Arten dieser Gruppe gemeinsam haben, ist das Fehlen einer Exine, einer Schutzschicht, um das Pollenkorn. Paul Knuth identifizierte 1906 zwei Arten der hydrophilen Bestäubung und Ernst Schwarzenbach fügte 1944 eine dritte hinzu. Knuth nannte seine beiden Gruppen Hyphydrogamie und die häufigere Ephydrogamie. Bei der Hyphydrogamie erfolgt die Bestäubung unterhalb der Wasseroberfläche, so dass die Pollenkörner in der Regel einen negativen Auftrieb haben. Bei Meerespflanzen, die diese Methode anwenden, sind die Narben in der Regel steif, während Süßwasserarten kleine und federartige Narben haben. Bei der Ephydrogamie findet die Bestäubung an der Wasseroberfläche statt, so dass die Pollenkörner eine geringe Dichte haben, um schwimmfähig zu sein, obwohl viele auch Flöße verwenden und hydrophob sind. Meeresblumen haben schwimmende fadenförmige Narben und sind möglicherweise an die Gezeiten angepasst, während Süßwasserarten Vertiefungen im Wasser bilden. Die dritte Kategorie, die von Schwarzenbach aufgestellt wurde, sind die Blumen, die den Pollen über dem Wasser transportieren, indem sie sich fortbewegen. Dies reicht von den Schwimmpflanzen (Lemnoideae) bis zu den staminaten Blüten (Vallisneria). Die meisten Arten dieser Gruppe haben trockene, kugelförmige Pollen, die sich manchmal zu größeren Massen zusammenlagern, und weibliche Blüten, die Vertiefungen im Wasser bilden; die Art des Transports variiert. ⓘ
Mechanismen
Blüten können durch zwei Mechanismen bestäubt werden: Fremdbestäubung und Selbstbestäubung. Kein Mechanismus ist unbestreitbar besser als der andere, da jeder seine Vor- und Nachteile hat. Je nach Lebensraum und ökologischer Nische nutzen die Pflanzen einen oder beide Mechanismen. ⓘ
Fremdbestäubung
Unter Fremdbestäubung versteht man die Bestäubung des Fruchtknotens durch Pollen einer anderen Pflanze der gleichen Art. Da die genetische Zusammensetzung der im Pollen der anderen Pflanze enthaltenen Spermien unterschiedlich ist, entsteht durch den Prozess der sexuellen Fortpflanzung eine neue, genetisch unterschiedliche Pflanze. Da jede neue Pflanze genetisch verschieden ist, unterscheiden sich die verschiedenen Pflanzen in ihren physiologischen und strukturellen Anpassungen, so dass die Population als Ganzes besser auf ungünstige Umwelteinflüsse vorbereitet ist. Die Fremdbestäubung erhöht daher das Überleben der Art und wird aus diesem Grund von den Blumen in der Regel bevorzugt. ⓘ
Selbstbestäubung
_flowers.jpg)
Unter Selbstbestäubung versteht man die Bestäubung des Fruchtknotens einer Blüte durch Pollen entweder von derselben Blüte oder einer anderen Blüte derselben Pflanze, was zur Bildung eines genetischen Klons durch ungeschlechtliche Fortpflanzung führt. Dies erhöht die Zuverlässigkeit der Samenproduktion, die Geschwindigkeit, mit der Samen produziert werden können, und senkt den Energiebedarf. Vor allem aber wird dadurch die genetische Variation eingeschränkt. Der Extremfall der Selbstbefruchtung, bei dem die Eizelle durch Pollen derselben Blume oder Pflanze befruchtet wird, tritt bei Blüten auf, die sich immer selbst befruchten, wie z. B. bei vielen Löwenzähnen. Einige Blumen sind selbstbefruchtend und haben Blüten, die sich nie öffnen, oder sie werden selbstbestäubt, bevor sich die Blüten öffnen; diese Blumen werden als kleistogam bezeichnet; dies ist beispielsweise bei vielen Arten der Gattung Viola der Fall. Umgekehrt gibt es bei vielen Pflanzenarten Möglichkeiten, die Selbstbestäubung und damit die Selbstbefruchtung zu verhindern. So können eingeschlechtliche männliche und weibliche Blüten an derselben Pflanze nicht zur gleichen Zeit erscheinen oder reifen, oder der Pollen derselben Pflanze ist nicht in der Lage, ihre Eizellen zu befruchten. Die letztgenannten Blütentypen, die chemische Barrieren gegen ihren eigenen Pollen haben, werden als selbstinkompatibel bezeichnet. Bei Clianthus puniceus (im Bild) wird die Selbstbestäubung strategisch als "Versicherungspolice" eingesetzt. Wenn ein Bestäuber, in diesem Fall ein Vogel, C. puniceus besucht, reibt er die Narbenhülle ab und ermöglicht es dem Pollen des Vogels, in die Narbe einzudringen. Wenn jedoch keine Bestäuber kommen, fällt die Narbenhülle auf natürliche Weise ab, so dass die eigenen Staubbeutel der Blume die Blume durch Selbstbestäubung bestäuben können. ⓘ
Allergien
Pollen tragen in hohem Maße zu Asthma und anderen Allergien der Atemwege bei, von denen insgesamt zwischen 10 und 50 % der Menschen weltweit betroffen sind. Diese Zahl scheint noch zu steigen, da der Temperaturanstieg infolge des Klimawandels dazu führt, dass die Pflanzen mehr Pollen produzieren, die auch mehr Allergien auslösen. Es ist jedoch schwierig, Pollen zu vermeiden, da sie sehr klein sind und in der Natur weit verbreitet sind. Die meisten Pollen, die Allergien auslösen, werden von durch den Wind verbreiteten Bestäubern wie Gräsern, Birken, Eichen und Ambrosia produziert; die Allergene in den Pollen sind Proteine, von denen man annimmt, dass sie für den Bestäubungsprozess notwendig sind. ⓘ
Befruchtung
_(1914)_(14590686978).jpg)
Die Befruchtung, auch Synagmie genannt, erfolgt im Anschluss an die Bestäubung, d. h. die Bewegung des Pollens vom Staubblatt zum Fruchtknoten. Sie umfasst sowohl die Plasmogamie, also die Verschmelzung der Protoplasten, als auch die Karyogamie, also die Verschmelzung der Zellkerne. Wenn der Pollen auf der Narbe der Blüte landet, beginnt er, einen Pollenschlauch zu bilden, der durch den Griffel in den Fruchtknoten läuft. Nachdem er in den mittleren Teil des Fruchtknotens eingedrungen ist, dringt er in den Eiapparat und in eine Synergide ein. An diesem Punkt platzt das Ende des Pollenschlauchs und setzt die beiden Samenzellen frei, von denen eine den Weg zur Eizelle findet, wobei sie auch ihre Zellmembran und einen Großteil ihres Protoplasmas verliert. Der Zellkern des Spermiums verschmilzt dann mit dem Kern der Eizelle, wodurch eine Zygote, eine diploide (zwei Kopien jedes Chromosoms) Zelle, entsteht. ⓘ
Während bei der Befruchtung nur die Plasmogamie, d. h. die Verschmelzung der gesamten Geschlechtszellen, stattfindet, kommt es bei den Angiospermen (Blütenpflanzen) zu einem Prozess, der als doppelte Befruchtung bezeichnet wird und sowohl Karyogamie als auch Plasmogamie beinhaltet. Bei der doppelten Befruchtung dringt die zweite Samenzelle anschließend ebenfalls in den Synergismus ein und verschmilzt mit den beiden polaren Kernen der Zentralzelle. Da alle drei Kerne haploid sind, entsteht ein großer Endospermkern, der triploid ist. ⓘ
Entwicklung des Samens

Nach der Bildung der Zygote beginnt diese durch Kern- und Zellteilungen, Mitose genannt, zu wachsen und wird schließlich zu einer kleinen Gruppe von Zellen. Ein Teil davon wird zum Embryo, der andere zum Suspensor, einer Struktur, die den Embryo in das Endosperm drückt und später nicht mehr nachweisbar ist. Zu diesem Zeitpunkt bilden sich auch zwei kleine Primordien, die später zum Keimblatt werden, das als Energiespeicher dient. Pflanzen, die aus einem dieser Keimblätter wachsen, werden Monokotyledonen genannt, während diejenigen, die aus zwei Keimblättern wachsen, Dikotyledonen sind. Das nächste Stadium wird als Torpedo-Stadium bezeichnet und umfasst das Wachstum mehrerer Schlüsselstrukturen, darunter die Keimwurzel (embryotische Wurzel), den Epikotyl (embryotischer Stamm) und den Hypokotyl (die Wurzel-Spross-Verbindung). In der letzten Phase entwickelt sich das Gefäßgewebe um den Samen. ⓘ
Entwicklung der Frucht
Der Fruchtknoten, in dem sich der Samen aus der Eizelle bildet, wächst zu einer Frucht heran. Alle anderen wichtigen Blütenteile sterben während dieser Entwicklung ab, darunter: Griffel, Narbe, Kelchblätter, Staubblätter und Blütenblätter. Die Frucht besteht aus drei Strukturen: dem Exokarp (äußere Schicht), dem Mesokarp (fleischiger Teil) und dem Endokarp (innerste Schicht), wobei die Fruchtwand als Perikarp bezeichnet wird. Größe, Form, Festigkeit und Dicke variieren von Frucht zu Frucht. Das liegt daran, dass sie direkt mit der Methode der Samenverbreitung zusammenhängt, d. h. mit dem Zweck der Frucht, die Verbreitung des Samens zu fördern oder zu ermöglichen und den Samen dabei zu schützen. ⓘ
Samenausbreitung
.jpg)


Nach der Bestäubung einer Blüte, der Befruchtung und schließlich der Entwicklung eines Samens und einer Frucht wird in der Regel ein Mechanismus eingesetzt, um die Frucht von der Pflanze weg zu verbreiten. Bei den Angiospermen (Blütenpflanzen) werden die Samen von der Pflanze weggestreut, um die Konkurrenz zwischen der Mutter- und den Tochterpflanzen zu vermeiden und die Besiedlung neuer Gebiete zu ermöglichen. Sie werden häufig in zwei Kategorien eingeteilt, obwohl viele Pflanzen dazwischen oder in eine oder mehrere dieser Kategorien fallen: ⓘ
Allochorie
Bei der Allochorie verwenden die Pflanzen einen externen Vektor oder Träger, um ihre Samen von ihnen weg zu transportieren. Diese können entweder biotisch (lebendig) sein, z. B. durch Vögel und Ameisen, oder abiotisch (nicht lebendig), z. B. durch den Wind oder das Wasser. ⓘ
Biotische Vektoren
Viele Pflanzen nutzen biotische Vektoren, um ihre Samen zu verbreiten. Diese Methode fällt unter den Oberbegriff Zoochorie, während sich die Endozoochorie, auch bekannt als Fruchtfresserei, speziell auf Pflanzen bezieht, die so angepasst sind, dass sie Früchte tragen, um Tiere anzulocken, die sie fressen. Nach dem Verzehr durchlaufen die Früchte in der Regel das Verdauungssystem des Tieres und werden von der Pflanze abgestoßen. Einige Samen sind speziell so angepasst, dass sie entweder im Magen von Tieren überleben oder sogar besser keimen, nachdem sie diese passiert haben. Sie können von Vögeln (Ornithochorie), Fledermäusen (Chiropterochorie), Nagetieren, Primaten, Ameisen (Myrmecochorie), vogelfreien Sauropsiden (Saurochorie), Säugetieren im Allgemeinen (Säugetieriochorie) und sogar Fischen gefressen werden. Ihre Früchte sind in der Regel fleischig, haben einen hohen Nährwert und enthalten möglicherweise chemische Lockstoffe als zusätzliche "Belohnung" für die Ausbreitungsorgane. Dies spiegelt sich morphologisch im Vorhandensein von mehr Fruchtfleisch, einem Arillus und manchmal einem Elaiosom (vor allem für Ameisen) wider, bei dem es sich um andere fleischige Strukturen handelt. Epizoochorie tritt bei Pflanzen auf, deren Samen so angepasst sind, dass sie sich an Tieren festhalten und auf diese Weise verbreitet werden können, wie bei vielen Arten der Gattung Acaena. Typischerweise haben die Samen dieser Pflanzen Haken oder eine zähflüssige Oberfläche, um sich leichter an Tieren festhalten zu können, darunter Vögel und Tiere mit Fell. Einige Pflanzen nutzen die Mimese, also die Nachahmung, um Tiere dazu zu bringen, die Samen zu verbreiten, und diese haben oft speziell angepasste Farben. Die letzte Art der Zoochorie wird als Synzoochorie bezeichnet, bei der die Samen weder verdaut noch unbeabsichtigt am Körper getragen werden, sondern von den Tieren absichtlich mitgeführt werden. Dies geschieht in der Regel im Maul oder im Schnabel des Tieres (Stomatochorie genannt), wie es bei vielen Vögeln und allen Ameisen der Fall ist.

.jpg)
Abiotische Vektoren
Bei der abiotischen Ausbreitung nutzen Pflanzen die Vektoren Wind, Wasser oder einen eigenen Mechanismus, um ihre Samen zu verbreiten. Bei der Anemochorie wird der Wind als Vektor genutzt, um die Pflanzensamen zu verbreiten. Da diese Samen mit dem Wind transportiert werden müssen, sind sie fast immer klein - manchmal sogar staubförmig -, haben ein großes Verhältnis von Oberfläche zu Volumen und werden in großer Zahl produziert - manchmal bis zu einer Million. Bei Pflanzen wie dem Steppengras wird der gesamte Trieb abgetrennt, damit die Samen mit dem Wind wegrollen können. Eine weitere häufige Anpassung sind Flügel, Federn oder ballonartige Strukturen, die es den Samen ermöglichen, länger in der Luft zu bleiben und somit weiter zu reisen. Bei der Hydrochorie sind die Pflanzen so angepasst, dass sie ihre Samen durch Wasser verbreiten können, weshalb sie in der Regel schwimmfähig sind und eine geringe relative Dichte im Verhältnis zum Wasser haben. Die Samen sind in der Regel morphologisch angepasst mit hydrophoben Oberflächen, geringer Größe, Haaren, Schleim, Öl und manchmal Lufträumen im Inneren der Samen. Diese Pflanzen lassen sich in drei Kategorien einteilen: solche, bei denen die Samen auf der Oberfläche von Wasserströmungen, unter der Oberfläche von Wasserströmungen und durch Regen, der auf einer Pflanze landet, verbreitet werden. ⓘ
Autochorie
Bei der Autochorie schaffen die Pflanzen ihre eigenen Vektoren, um die Samen von sich weg zu transportieren. Dies geschieht in der Regel dadurch, dass die Früchte explodieren und die Samen ballistisch weggeschleudert werden, wie z. B. bei Hura crepitans, oder manchmal auch durch die Bildung von kriechenden Diasporen. Wegen der relativ geringen Entfernungen, über die diese Methoden ihre Samen verbreiten können, werden sie oft mit einem externen Vektor gepaart. ⓘ
Entwicklung
Landpflanzen gibt es zwar schon seit etwa 425 Millionen Jahren, aber die ersten vermehrten sich durch eine einfache Anpassung ihrer aquatischen Gegenstücke: Sporen. Im Meer können Pflanzen - und einige Tiere - einfach genetische Klone von sich selbst ausstreuen, die dann davonschwimmen und woanders wachsen. Auf diese Weise vermehrten sich die frühen Pflanzen. Doch schon bald entwickelten die Pflanzen Methoden zum Schutz dieser Kopien, um sie vor Austrocknung und anderen Schäden zu schützen, die an Land noch wahrscheinlicher sind als im Meer. Der Schutz wurde zum Samen, obwohl sich die Blüte noch nicht entwickelt hatte. Zu den frühen samentragenden Pflanzen gehören der Ginkgo und die Koniferen. ⓘ

Mehrere Gruppen von ausgestorbenen Gymnospermen, insbesondere Samenfarne, wurden als Vorfahren der Blütenpflanzen vorgeschlagen, aber es gibt keine kontinuierlichen fossilen Beweise, die genau zeigen, wie sich Blumen entwickelt haben. Das scheinbar plötzliche Auftauchen relativ moderner Blumen im Fossilbericht stellte ein solches Problem für die Evolutionstheorie dar, dass Charles Darwin es ein "abscheuliches Rätsel" nannte. ⓘ
Kürzlich entdeckte Fossilien von Angiospermen wie Archaefructus sowie weitere Entdeckungen von fossilen Gymnospermen lassen vermuten, dass die Merkmale der Angiospermen in mehreren Schritten erworben wurden. Ein frühes Fossil einer Blütenpflanze, Archaefructus liaoningensis aus China, ist etwa 125 Millionen Jahre alt. Noch älter ist der 125-130 Millionen Jahre alte Archaefructus sinensis aus China. Im Jahr 2015 wurde eine Pflanze (die 130 Millionen Jahre alte Montsechia vidalii, die in Spanien entdeckt wurde) auf ein Alter von 130 Millionen Jahren geschätzt. Im Jahr 2018 berichteten Wissenschaftler, dass die frühesten Blüten vor etwa 180 Millionen Jahren entstanden. ⓘ
_fragment.jpg)
Jüngste DNA-Analysen (molekulare Systematik) zeigen, dass Amborella trichopoda, die auf der Pazifikinsel Neukaledonien vorkommt, die einzige Art in der Schwestergruppe der übrigen Blütenpflanzen ist, und morphologische Untersuchungen deuten darauf hin, dass sie Merkmale aufweist, die für die frühesten Blütenpflanzen charakteristisch gewesen sein könnten. ⓘ
Neben den eindeutigen Beweisen für Blüten in oder kurz vor der Kreidezeit gibt es einige Indizien, die auf Blüten vor 250 Millionen Jahren hindeuten. Eine Chemikalie, die von Pflanzen zur Verteidigung ihrer Blüten verwendet wird, Oleanan, wurde in fossilen Pflanzen aus dieser Zeit nachgewiesen, darunter auch in Gigantopteriden, die sich zu dieser Zeit entwickelten und viele Merkmale moderner Blütenpflanzen aufweisen, obwohl sie selbst nicht als Blütenpflanzen bekannt sind, da nur ihre Stängel und Stacheln im Detail erhalten sind; eines der frühesten Beispiele für Versteinerung. ⓘ
Die Ähnlichkeit in der Blatt- und Stängelstruktur kann sehr wichtig sein, denn Blumen sind genetisch gesehen nur eine Anpassung der normalen Blatt- und Stängelkomponenten von Pflanzen, eine Kombination von Genen, die normalerweise für die Bildung neuer Triebe verantwortlich sind. Es wird angenommen, dass die primitivsten Blüten eine unterschiedliche Anzahl von Blütenteilen hatten, die oft voneinander getrennt waren (aber in Kontakt miteinander standen). Die Blüten wuchsen in der Regel spiralförmig, waren zweigeschlechtlich (bei Pflanzen bedeutet dies, dass sowohl männliche als auch weibliche Teile auf derselben Blüte vorhanden waren) und wurden vom Fruchtknoten (dem weiblichen Teil) dominiert. Mit fortschreitender Entwicklung der Blüten entwickelten sich einige Varianten, bei denen die Teile miteinander verschmolzen sind, mit einer viel spezifischeren Anzahl und Gestaltung und entweder mit spezifischen Geschlechtern pro Blüte oder Pflanze oder zumindest mit einem "untergeordneten Fruchtknoten". ⓘ
Allgemein wird davon ausgegangen, dass die Funktion der Blüten von Anfang an darin bestand, Tiere in den Fortpflanzungsprozess einzubeziehen. Die Pollen können ohne leuchtende Farben und auffällige Formen gestreut werden, die somit eine Belastung für die Ressourcen der Pflanze darstellen würden, sofern sie nicht einen anderen Nutzen bringen. Ein möglicher Grund für das plötzliche, voll entwickelte Auftreten von Blüten ist, dass sie sich in einer isolierten Umgebung wie einer Insel oder einer Inselkette entwickelt haben, wo die Pflanzen, die sie tragen, eine hochspezialisierte Beziehung zu einem bestimmten Tier (z. B. einer Wespe) entwickeln konnten, wie es heute bei vielen Inselarten der Fall ist. Diese symbiotische Beziehung, bei der eine hypothetische Wespe Pollen von einer Pflanze zur anderen trägt, ähnlich wie es heute Feigenwespen tun, könnte schließlich dazu geführt haben, dass sowohl die Pflanze(n) als auch ihre Partner einen hohen Grad an Spezialisierung entwickelt haben. Man geht davon aus, dass die Inselgenetik eine häufige Quelle der Speziation ist, insbesondere wenn es um radikale Anpassungen geht, die anscheinend minderwertige Übergangsformen erforderten. Das Beispiel der Wespe ist nicht zufällig: Bienen, die sich offenbar speziell für symbiotische Pflanzenbeziehungen entwickelt haben, stammen von Wespen ab. ⓘ
Ebenso stammen die meisten Früchte, die bei der pflanzlichen Fortpflanzung verwendet werden, von der Vergrößerung von Teilen der Blüte ab. Diese Frucht ist häufig ein Werkzeug, das von Tieren abhängt, die es fressen wollen und daher die darin enthaltenen Samen verstreuen. ⓘ
Während viele dieser symbiotischen Beziehungen zu zerbrechlich sind, um im Wettbewerb mit den Organismen auf dem Festland zu überleben, erwiesen sich die Blüten als ungewöhnlich wirksame Produktionsmittel, die sich (unabhängig von ihrem tatsächlichen Ursprung) zur vorherrschenden Form des Pflanzenlebens an Land ausbreiteten. ⓘ
Die Evolution der Blumen setzt sich bis heute fort; die modernen Blumen wurden vom Menschen so stark beeinflusst, dass viele von ihnen in der Natur nicht bestäubt werden können. Viele moderne, domestizierte Blumen waren früher einfache Unkräuter, die nur dann sprossen, wenn der Boden aufgewühlt wurde. Einige von ihnen neigten dazu, mit der menschlichen Ernte zu wachsen, und die schönsten wurden wegen ihrer Schönheit nicht gepflückt, wodurch sich eine Abhängigkeit von der menschlichen Zuneigung und eine besondere Anpassung an diese entwickelte. ⓘ
Farbe
thumb|Reflektionsspektren für die Blüten verschiedener Rosensorten. Eine rote Rose absorbiert etwa 99,7 % des Lichts in einem breiten Bereich unterhalb der roten Wellenlängen des Spektrums, was zu einem außergewöhnlich reinen Rot führt. Eine gelbe Rose reflektiert etwa 5 % des blauen Lichts, wodurch ein ungesättigtes Gelb entsteht (ein Gelb mit einem gewissen Weißanteil). Viele blühende Pflanzen reflektieren so viel Licht wie möglich im Bereich der sichtbaren Wellenlängen des Bestäubers, den die Pflanze anlocken soll. Blüten, die das gesamte Spektrum des sichtbaren Lichts reflektieren, werden von einem menschlichen Betrachter im Allgemeinen als weiß wahrgenommen. Ein wichtiges Merkmal weißer Blüten ist, dass sie das gesamte sichtbare Spektrum gleichmäßig reflektieren. Viele Blütenpflanzen verwenden Weiß, um Bestäuber anzulocken, aber auch die Verwendung von Farbe ist weit verbreitet (sogar innerhalb derselben Art). Mit Hilfe der Farbe kann eine blühende Pflanze den Bestäuber, den sie anlocken möchte, genauer bestimmen. Das von der menschlichen Farbreproduktionstechnologie (CMYK) verwendete Farbmodell beruht auf der Modulation von Pigmenten, die das Spektrum in breite Absorptionsbereiche unterteilen. Blühende Pflanzen hingegen sind in der Lage, die Wellenlänge des Übergangspunkts zwischen Absorption und Reflexion zu verschieben. Wenn man davon ausgeht, dass das visuelle System der meisten Bestäuber das sichtbare Spektrum als kreisförmig ansieht, kann man sagen, dass blühende Pflanzen Farbe erzeugen, indem sie das Licht in einem Bereich des Spektrums absorbieren und in dem anderen Bereich reflektieren. Bei CMYK wird die Farbe in Abhängigkeit von der Amplitude der breiten Absorptionsbereiche erzeugt. Blühende Pflanzen hingegen erzeugen Farbe, indem sie die Frequenz (oder vielmehr die Wellenlänge) des reflektierten Lichts verändern. Die meisten Blüten absorbieren Licht aus dem blauen bis gelben Bereich des Spektrums und reflektieren Licht aus dem grünen bis roten Bereich des Spektrums. Bei vielen blühenden Pflanzenarten ist es der Übergangspunkt, der die Farbe charakterisiert, die sie erzeugen. Die Farbe kann durch Verschiebung des Übergangspunkts zwischen Absorption und Reflexion moduliert werden, und auf diese Weise kann eine Blütenpflanze angeben, welchen Bestäuber sie anlocken möchte. Einige Blütenpflanzen haben auch eine begrenzte Fähigkeit, die Absorptionsbereiche zu modulieren. Dies ist in der Regel nicht so präzise wie die Kontrolle über die Wellenlänge. Der menschliche Beobachter nimmt dies als Sättigungsgrad wahr (der Weißanteil der Farbe). ⓘ
Klassische Taxonomie
.png)
In der Pflanzentaxonomie, die sich mit der Klassifizierung und Identifizierung von Pflanzen befasst, wird die Morphologie der Pflanzenblüten ausgiebig genutzt - und das schon seit Tausenden von Jahren. Obwohl die Geschichte der Pflanzentaxonomie mindestens bis etwa 300 v. Chr. mit den Schriften von Theophrastus zurückreicht, beruht die Grundlage der modernen Wissenschaft auf Arbeiten aus dem 18. und 19. ⓘ
Carl Linnaeus (1707-1778) war ein schwedischer Botaniker, der den größten Teil seines Berufslebens als Professor für Naturgeschichte verbrachte. In seinem bahnbrechenden Buch Species Plantarum aus dem Jahr 1757 legte er sein Klassifizierungssystem sowie das Konzept der binomischen Nomenklatur dar, das auch heute noch weltweit verwendet wird. Er ermittelte 24 Klassen, die hauptsächlich auf der Anzahl, der Länge und der Verbindung der Staubgefäße basieren. Die ersten zehn Klassen richten sich direkt nach der Anzahl der Staubgefäße (Octandria haben 8 Staubgefäße usw.), während die elfte Klasse 11-20 Staubgefäße und die zwölfte und dreizehnte Klasse 20 Staubgefäße haben; sie unterscheiden sich nur durch ihren Ansatzpunkt. Die nächsten fünf Klassen befassen sich mit der Länge der Staubgefäße und die letzten fünf mit der Art der Fortpflanzungsfähigkeit der Pflanze, dem Ort, an dem die Staubgefäße wachsen, und ob die Blüte verdeckt ist oder überhaupt existiert (wie bei Farnen). Obwohl diese Klassifizierungsmethode künstlich war, wurde sie in den folgenden sieben Jahrzehnten häufig verwendet, bevor sie durch das System eines anderen Botanikers ersetzt wurde. ⓘ
Antoine Laurent de Jussieu (1748-1836) war ein französischer Botaniker, der 1787 in seinem Werk Genera plantarum: secundum ordines naturales disposita eine neue Methode zur Klassifizierung von Pflanzen vorstellte, die auf natürlichen Merkmalen beruhte. Die Pflanzen wurden nach der Anzahl der Keimblätter (sofern vorhanden) und der Lage der Staubgefäße eingeteilt. Das nächste bedeutende Klassifizierungssystem wurde im späten 19. Jahrhundert von den Botanikern Joseph Dalton Hooker (1817-1911) und George Bentham (1800-1884) entwickelt. Sie bauten auf den früheren Arbeiten von de Jussieu und Augustin Pyramus de Candolle auf und entwickelten ein System, das noch immer in vielen Herbarien der Welt verwendet wird. Die Pflanzen wurden auf der obersten Ebene nach der Anzahl der Keimblätter und der Art der Blüten unterteilt, bevor sie in Ordnungen (Familien), Gattungen und Arten eingeteilt wurden. Dieses Klassifizierungssystem wurde in den Genera plantarum in drei Bänden zwischen 1862 und 1883 veröffentlicht. Es ist das am meisten geschätzte System und gilt in manchen Kreisen als das beste Klassifizierungssystem". ⓘ
Im Zuge der Entwicklung des wissenschaftlichen Denkens nach Darwins Über die Entstehung der Arten haben viele Botaniker phylogenetische Methoden angewandt, und der Einsatz von genetischer Sequenzierung, Zytologie und Palynologie ist zunehmend üblich geworden. Dennoch bilden morphologische Merkmale wie die Beschaffenheit der Blüte und des Blütenstandes nach wie vor die Grundlage der Pflanzentaxonomie. ⓘ
Symbolik

Viele Blumen haben in der westlichen Kultur wichtige symbolische Bedeutungen. Die Praxis der Zuordnung von Bedeutungen zu Blumen ist als Floriografie bekannt. Einige der gängigsten Beispiele sind:
- Rote Rosen werden als Symbol für Liebe, Schönheit und Leidenschaft verschenkt.
- Mohnblumen sind ein Symbol des Trostes in Zeiten des Todes. Im Vereinigten Königreich, in Neuseeland, Australien und Kanada werden rote Mohnblumen getragen, um den im Krieg gefallenen Soldaten zu gedenken.
- Schwertlilien/Lilien werden bei Beerdigungen als Symbol für die "Auferstehung/das Leben" verwendet. Sie wird auch mit den Sternen (Sonne) und ihren blühenden/leuchtenden Blütenblättern in Verbindung gebracht.
- Gänseblümchen sind ein Symbol der Unschuld. ⓘ

Aufgrund ihrer vielfältigen und farbenfrohen Erscheinung sind Blumen seit langem auch ein beliebtes Thema für bildende Künstler. Einige der berühmtesten Gemälde bekannter Maler zeigen Blumen, wie die Sonnenblumen-Serie von Van Gogh oder die Seerosen von Monet. Blumen werden auch getrocknet, gefriergetrocknet und gepresst, um dauerhafte, dreidimensionale florale Kunstwerke zu schaffen. ⓘ
Blumen in der Kunst stehen auch für die weiblichen Genitalien, wie in den Werken von Künstlern wie Georgia O'Keeffe, Imogen Cunningham, Veronica Ruiz de Velasco und Judy Chicago zu sehen ist, und zwar in der klassischen asiatischen und westlichen Kunst. Viele Kulturen auf der ganzen Welt neigen dazu, Blumen mit Weiblichkeit zu assoziieren. ⓘ
Die große Vielfalt der zarten und schönen Blumen hat zahlreiche Dichter inspiriert, vor allem in der Romantik des 18. bis 19. Berühmte Beispiele sind William Wordsworths I Wandered Lonely as a Cloud und William Blakes Ah! Sun-Flower. ⓘ
Ihre Symbolik in Träumen wurde ebenfalls diskutiert, wobei mögliche Interpretationen "blühendes Potenzial" sind. ⓘ
Die römische Göttin der Blumen, der Gärten und des Frühlings ist Flora. Die griechische Göttin des Frühlings, der Blumen und der Natur ist Chloris. ⓘ
In der hinduistischen Mythologie haben Blumen einen hohen Stellenwert. Vishnu, einer der drei Hauptgötter im hinduistischen System, wird oft aufrecht auf einer Lotusblume stehend dargestellt. Abgesehen von der Assoziation mit Vishnu kommt dem Lotus in der hinduistischen Tradition auch eine spirituelle Bedeutung zu. So kommt er beispielsweise in den hinduistischen Schöpfungsgeschichten vor. ⓘ
Menschliche Verwendung

Die Geschichte zeigt, dass Blumen seit Jahrtausenden von den Menschen zu verschiedenen Zwecken verwendet wurden. Ein frühes Beispiel dafür ist das alte Ägypten vor etwa 4.500 Jahren, wo Blumen als Haarschmuck für Frauen verwendet wurden. Blumen haben auch immer wieder die Kunst inspiriert, wie in Monets Seerosen oder William Wordsworths Gedicht über Narzissen mit dem Titel: "I Wandered Lonely as a Cloud". ⓘ
In der Neuzeit haben die Menschen nach Möglichkeiten gesucht, Blumen und blühende Pflanzen zu züchten, zu kaufen, zu tragen oder sich anderweitig mit ihnen zu umgeben, teilweise wegen ihres angenehmen Aussehens und Geruchs. Überall auf der Welt verwenden die Menschen Blumen, um wichtige Ereignisse in ihrem Leben zu markieren:
- Bei Neugeburten oder Taufen
- als Ansteckblume oder Boutonniere bei gesellschaftlichen Anlässen oder zu Feiertagen
- als Zeichen der Liebe oder Wertschätzung
- als Hochzeitsblumen für die Hochzeitsgesellschaft und als Dekoration für den Saal
- als verschönernde Dekoration in den eigenen vier Wänden
- als Erinnerungsgeschenk für Abschiedsfeiern, Willkommensfeiern und "Denk an dich"-Geschenke
- Für Trauerflor und Beileidsbekundungen für Trauernde
- Für Gottesdienste. Im Christentum schmücken Blumen im Altarraum oft die Kirchen. In der hinduistischen Kultur bringen die Anhänger üblicherweise Blumen als Geschenk zu Tempeln mit. ⓘ

Blumen wie Jasmin werden in China seit Jahrhunderten als Ersatz für den traditionellen Tee verwendet. In jüngster Zeit gewinnen viele andere Kräuter und Blumen, die traditionell in der ganzen Welt verwendet werden, an Bedeutung, um eine Reihe von Blütentees zuzubereiten. ⓘ
Die Menschen züchten daher Blumen rund um ihr Haus, widmen einen Teil ihres Wohnraums einem Blumengarten, pflücken Wildblumen oder kaufen kommerziell gezüchtete Blumen bei Blumenhändlern. ⓘ

Blumen liefern zwar weniger Nahrung als andere wichtige Pflanzenteile (Samen, Früchte, Wurzeln, Stängel und Blätter), aber dennoch einige wichtige Gemüse und Gewürze. Zu den Blumengemüsen gehören Brokkoli, Blumenkohl und Artischocken. Das teuerste Gewürz, Safran, besteht aus den getrockneten Blütennarben eines Krokus. Andere Blütengewürze sind Nelken und Kapern. Hopfenblüten werden zum Aromatisieren von Bier verwendet. Ringelblumenblüten werden an Hühner verfüttert, um deren Eigelb eine goldgelbe Farbe zu verleihen, die von den Verbrauchern als begehrenswerter empfunden wird; getrocknete und gemahlene Ringelblumenblüten werden auch in der georgischen Küche als Gewürz und Farbstoff verwendet. Blüten des Löwenzahns und des Holunders werden häufig zu Wein verarbeitet. Bienenpollen, von Bienen gesammelter Blütenstaub, wird von manchen Menschen als gesundes Nahrungsmittel angesehen. Honig besteht aus von Bienen verarbeiteten Blütennektar und wird oft nach der Art der Blume benannt, z. B. Orangenblütenhonig, Kleehonig und Tupelohonig. ⓘ
Hunderte von frischen Blüten sind essbar, aber nur wenige werden als Lebensmittel vermarktet. Sie werden oft als Beilage zu Salaten gereicht. Kürbisblüten werden in Paniermehl getaucht und gebraten. Zu den essbaren Blumen gehören Kapuzinerkresse, Chrysanthemen, Nelken, Katzenschwanz, japanisches Geißblatt, Zichorie, Kornblume, Canna und Sonnenblume. Essbare Blumen wie Gänseblümchen, Rosen und Veilchen werden manchmal kandiert. ⓘ
Blumen wie Chrysantheme, Rose, Jasmin, japanisches Geißblatt und Kamille werden wegen ihres Duftes und ihrer medizinischen Eigenschaften als Tisane verwendet, entweder mit Tee gemischt oder pur. ⓘ
Blumen werden seit prähistorischen Zeiten bei Begräbnisritualen verwendet: Spuren von Blütenstaub wurden auf dem Grab einer Frau in der Höhle von El Miron in Spanien gefunden. Viele Kulturen stellen eine Verbindung zwischen Blumen und Leben und Tod her, und aufgrund ihrer jahreszeitlichen Wiederkehr suggerieren Blumen auch Wiedergeburt, was erklären mag, warum viele Menschen Blumen auf Gräber legen. Wie in Euripides' Stück Die phönizischen Frauen beschrieben, setzten die alten Griechen den Verstorbenen einen Blumenkranz auf den Kopf und bedeckten ihre Gräber mit Kränzen und Blütenblättern. Blumen waren bei altägyptischen Bestattungen weit verbreitet, und die Mexikaner verwenden bis heute Blumen bei ihren Feiern zum Tag der Toten, so wie es ihre aztekischen Vorfahren taten.

Schenken

Die Tradition des Blumenschenkens geht auf prähistorische Zeiten zurück, als Blumen oft medizinische und pflanzliche Eigenschaften besaßen. Archäologen fanden in mehreren Gräbern Überreste von Blütenblättern. Blumen wurden zunächst als Opfer- und Grabbeigaben verwendet. Die alten Ägypter und später die Griechen und Römer verwendeten Blumen. In Ägypten wurden Grabbeigaben aus der Zeit um 1540 v. Chr. gefunden, auf denen roter Mohn, gelber Araun, Kornblumen und Lilien abgebildet waren. Aufzeichnungen über das Schenken von Blumen finden sich in chinesischen Schriften und ägyptischen Hieroglyphen sowie in der griechischen und römischen Mythologie. Der Brauch, Blumen zu schenken, blühte im Mittelalter auf, als Paare ihre Zuneigung durch Blumen zeigten. ⓘ
Die Tradition des Blumenschenkens existiert in vielen Formen. Sie ist ein wichtiger Teil der russischen Kultur und Folklore. Es ist üblich, dass Schüler ihren Lehrern Blumen schenken. Gelbe Blumen in einer romantischen Beziehung zu verschenken, bedeutet in Russland eine Trennung. Heutzutage werden Blumen oft in Form eines Blumenstraußes verschenkt. ⓘ
Blütenbildung
Nachdem durch die Heterochroniegene ein Meristem vom vegetativen in ein generatives Meristem umgewandelt wurde, erfolgt durch eine Vielzahl von interagierenden Regulator-Genen die eigentliche Blütenbildung. ⓘ
Symmetriegene
Sogenannte Symmetrie- oder Katastergene legen die Struktur der Blüte und ihre Symmetrieebenen fest, sowie die Anzahl der Blüten und Blütenorgane. Mutationen dieser Gene verändern die Architektur der Blüte, die Gene werden vielfach nach den durch solche Mutationen veränderten Phänotypen benannt: Mutationen der Gene Cycloidea (CYC) oder Dichotoma (DICH) beim Löwenmäulchen erzeugen radiärsymmetrische Blüten anstelle der üblichen zygomorphen. Diese Mutationen treten auch natürlich auf und werden Pelorie genannt. Die Mutation des Gens Cauliflower (CAL) bei Arabidopsis und beim Brokkoli erzeugt viele Verzweigungen im Blütenstand ohne funktionsfähige Blüten. CAL hemmt das Gen TFL1, das die Verzweigung im Blütenstand fördert und ein Spross-Identitäts-Gen ist. tfl1-Mutanten bilden dementsprechend wenig- bis einblütige Blütenstände in Arabidopsis. ⓘ
Vielfalt des ABC-Modells

Vergleichende Untersuchungen bei einer Reihe von Pflanzenarten haben ergeben, dass das ABC-Modell in der oben beschriebenen Form nur für die Eudikotylen gelten dürfte. Besonders bei den basalen Gruppen der Angiospermen gibt es verschiedene Ausprägungen. Bei Teichrosen (Nuphar) gibt es keine scharfen Grenzen der Genexpression (“fading borders”), sodass es eine Übergangsreihe von kelchähnlichen Tepalen über kronblattähnliche Tepalen zu Staminodien zu Staubblättern zu Fruchtblättern gibt. ⓘ
Die Tulpe bildet eine einheitliche Blütenhülle in zwei Kreisen aus. Hier werden in beiden Kreisen B-Klasse-Gene exprimiert. ⓘ
Die Blüten der Süßgräser sind sehr stark abgeleitet. Dennoch zeigen sie das ABC-Modell, wobei hier die Vorspelze (Palea) durch A und die Schwellkörper (Lodiculae) durch A und B gekennzeichnet sind, also homolog zu Kelch und Krone. ⓘ
Der Ampfer (Rumex) bildet eine einheitliche, aber reduzierte und unauffällige Blütenhülle. Hier ist die B-Klasse auf die Staubblätter beschränkt, sodass beide Blütenhüllkreise nur durch A definiert werden. ⓘ
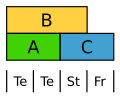
Abgewandeltes ABC-Modell der Tulpe. Te Tepalen
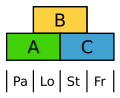
ABC-Modell beim Reis. Pa Vorspelze, Lo Schwellkörper
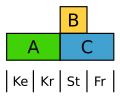
Abgewandeltes ABC-Modell beim Ampfer ⓘ



Inzwischen wird jedoch die Allgemeingültigkeit der A-Klasse als Blütenorganidentitätsgene angezweifelt. A-Klasse-Gene spielen, wie oben ausgeführt, auch eine Rolle bei der Festlegung der Blüten-Meristem-Identität. Nachgewiesen werden konnten sie in der Bildung der Blütenorgane bis jetzt nur bei Arabidopsis und sie dürften nur in der näheren Verwandtschaft von Bedeutung sein. Bei Antirrhinum reichen die BC-Gene zur Blütenbildung. Inwiefern dies für andere Angiospermen-Gruppen gilt, ist offen. ⓘ
Anthese
Der Vorgang des Blühens wird als Anthese (von griechisch anthēsis „Blüte“) bezeichnet. Es ist der Entwicklungsabschnitt der Blütenorgane vom Ende des Knospenzustandes bzw. Beginn der Knospenentfaltung bis zum Beginn des Verblühens. ⓘ
Blütenöffnung
Vor dem Öffnen bildet die Blüte eine Knospe ähnlich den Überwinterungsknospen. Dabei dienen die Kelchblätter als Schutz. Die Blütenöffnung gehört zu den Pflanzenbewegungen. Die Öffnung der Blütenknospe kann durch verschiedene Mechanismen erfolgen:
- Reversible Akkumulation von Ionen. Dies wurde bis jetzt nur bei dem Kochschen Enzian (Gentiana acaulis) eindeutig nachgewiesen.
- Programmierter Zelltod in definierten Bereichen der Blütenblätter.
- Wasserverlust am Tag und Wiederauffüllen während der Nacht. Dies tritt etwa bei Silene saxifraga auf, die nachtblühend ist und deren Kronblätter sich am Tag infolge Wasserverlustes einrollen.
- Differenzielles Wachstum der Außen- und Innenseite der Blütenblätter. Bei der Tulpe hat die Innenseite der Blütenblätter ein um 10 °C niedrigeres Wachstumsoptimum als die Außenseite, was zu einem Öffnen am Morgen und dem Schließen am Abend führt. ⓘ
Ein wichtiger Faktor für das Ausbreiten der Kronblätter ist die Erhöhung des Turgors, der meist durch eine Erhöhung der Zuckerkonzentration infolge Abbaus hochmolekularer Kohlenhydrate erfolgt (Stärke etwa bei Rosen, Fructane etwa bei Taglilien). Damit einher geht eine Expansion der Zellwand. Die Angaben über die Rolle von Pflanzenhormonen sind in der Literatur widersprüchlich. ⓘ
Das Öffnen der Blüten wird durch externe Faktoren ausgelöst. Bei nachtblühenden Arten ist häufig die erhöhte Luftfeuchtigkeit am Abend der Auslöser. Auf Temperaturerhöhung reagieren besonders Vorfrühlingsblüher, zum Beispiel das Schneeglöckchen (Galanthus nivalis) oder Krokusse (Crocus spp.). Eine dritte Gruppe reagiert auf Licht, wie das Gänseblümchen (Bellis perennis). Das Öffnen und Schließen langlebiger Blüten bei Tag beziehungsweise Nacht geschieht durch die gleichen Mechanismen und unterliegt einer endogenen Rhythmik. Die zellphysiologischen und besonders die genetischen Hintergründe der Blütenöffnung und -schließung sind bis jetzt kaum bekannt. ⓘ
Das Schließen der Blüten kann durch differenzielles Wachstum oder durch reversible Turgoränderungen geschehen. In diesen Fällen ist ein wiederholtes Öffnen und Schließen möglich. Turgorverlust durch Seneszenz führt zur permanenten Schließung der Blüte. ⓘ
Seneszenz
Die Blüten werden als Sexualorgane in den Pflanzen stets neu gebildet, im Gegensatz zu denen der Tiere. Die Lebensdauer ist genau abgemessen, da Blüten große Ressourcen verbrauchen und die Narbe auch ein wesentlicher Eintrittspunkt für Krankheitserreger ist. Bereits bestäubte Blüten würden auch unnötigerweise mit nicht bestäubten um Bestäuber konkurrieren. ⓘ
Ein wichtiger Auslöser für Seneszenz (Alterung) ist die Bestäubung mit Pollen. Diese verkürzt in den meisten Arten die Lebensdauer der Blüte beträchtlich. In etlichen Pflanzen löst die Bestäubung die Bildung des Pflanzenhormons Ethylen aus, welches wiederum die Seneszenz der Kronblätter auslöst. Andere Arten sind jedoch unempfindlich gegen Ethylen, ihr Seneszenz-Mechanismus ist unbekannt. Auf Organ-Ebene sterben nach der Bestäubung Krone, Staubgefäße und der Griffel ab, während sich der Fruchtknoten zur Frucht weiterentwickelt. Das Absterben geht mit einer Remobilisierung der Inhaltsstoffe einher, ähnlich der Seneszenz der Blätter. ⓘ
Evolution
Molekularbiologisch begründete Theorien
Die ältesten Angiospermenblüten waren zwittrig und hatten ein undifferenziertes Perianth in Spiralstellung oder mehr als zwei Wirteln. ⓘ
Außerhalb der Samenpflanzen gibt es keine Blütenorgan-Identitätsgene oder Orthologe davon. In den Gymnospermen, die nur eingeschlechtige Blüten bilden, gibt es die Orthologe der Klasse B und C. Ihre Expression gleicht der in Angiospermen: C wird in allen reproduktiven Organen ausgebildet, B in den männlichen Blüten. Basierend auf diesen Erkenntnissen, wurden mehrere Hypothesen zur Entstehung der zwittrigen Angiospermen-Blüte aufgestellt:
- Nach der “out of male”-Hypothese von Theissen et al. 2002 bildeten die männlichen Blütenzapfen durch eine Reduktion der B-Klassen-Expression im oberen Zapfenbereich weibliche Organe. Nach der “out of female” Hypothese bildeten sich die männlichen Organe am unteren Ende der weiblichen Zapfen. Eine Blütenhülle entstand nach diesen beiden Modellen erst nach der Zwittrigkeit.
- Ebenfalls von männlichen Blüten geht die “mostly male” Hypothese aus, zuerst vorgeschlagen von Frohlich und Parker 2000. Sie entdeckten, dass es in Gymnospermen das Leafy-Gen in zwei Kopien vorkommt (Paraloge). Leafy spezifiziert männliche, Needly weibliche Blüten. Needly kommt in allen Gymnospermen außer Gnetum vor, aber nicht in Angiospermen. Die Theorie besagt nun, dass durch Deaktivierung des Needly-Gens die männlichen Zapfen zwittrig wurden. ⓘ
Die Evolution der Angiospermen-Blüte fand in vier Schlüsselereignissen statt:
- Evolution der zwittrigen Blütenachse
- Evolution der gestauchten Blütenachse und die Begrenzung des Wachstums: Dies geschah durch C-Klasse-Gene, die Wuschel, das Meristem-Erhaltungsgen, unterdrücken.
- Evolution einer petaloiden Blütenhülle
- Evolution des klassischen zweikreisigen, zweiteiligen Perianths der Eudikotylen aus Kelch und Krone. ⓘ
Soltis et al. (2007) diskutieren die Hypothese, dass an der Basis der Angiospermen nicht das ABC-Modell wie in Arabidopsis stand, sondern ein System mit unscharfen Grenzen wie bei der Teichrose. Von diesem Grundmodell können dann einfach durch verschiedene Grenzschärfungen das ABC-Modell wie auch die abgewandelte Formen wie bei der Tulpe entstanden sein. ⓘ