Blütenstand


Ein Blütenstand ist eine Gruppe oder ein Büschel von Blüten, die an einem Stängel angeordnet sind, der aus einem Hauptast oder einer komplizierten Anordnung von Zweigen besteht. Morphologisch gesehen handelt es sich um den veränderten Teil des Sprosses von Samenpflanzen, in dem sich die Blüten an der Achse der Pflanze bilden. Die Modifikationen können die Länge und die Art der Internodien und der Phyllotaxis betreffen, sowie Variationen in den Proportionen, Verdichtungen, Anschwellungen, Verwachsungen, Konnationen und Reduktionen von Haupt- und Nebenachsen. Man kann einen Blütenstand auch als den reproduktiven Teil einer Pflanze definieren, der ein Bündel von Blüten in einem bestimmten Muster trägt. ⓘ
Der Stiel, der den gesamten Blütenstand trägt, wird als Blütenstiel bezeichnet. Die Hauptachse (fälschlicherweise als Hauptstamm bezeichnet) oberhalb des Blütenstiels, die die Blüten oder Nebenzweige trägt, wird als Spindel bezeichnet. Der Stiel jeder Blüte des Blütenstandes wird als Blütenstiel bezeichnet. Eine Blüte, die nicht Teil eines Blütenstandes ist, wird als Einzelblüte bezeichnet, und ihr Stiel wird ebenfalls als Blütenstiel bezeichnet. Jede Blüte in einem Blütenstand kann als Blüte bezeichnet werden, vor allem wenn die einzelnen Blüten besonders klein sind und in einer dichten Traube stehen, wie z. B. bei einem Pseudanthium. Das Fruchtstadium eines Blütenstandes wird als Infrukteszenz bezeichnet. Die Blütenstände können einfach (einzeln) oder komplex (Rispe) sein. Die Rispe kann eine von mehreren Arten sein, einschließlich einfach, zusammengesetzt, Dolde, Ähre oder Traube. ⓘ
Allgemeine Merkmale
Blütenstände werden durch viele verschiedene Merkmale beschrieben, z. B. durch die Anordnung der Blüten am Blütenstiel, die Reihenfolge der Blüte und die Gruppierung der verschiedenen Blütenbüschel. Bei diesen Begriffen handelt es sich um allgemeine Darstellungen, da Pflanzen in der Natur eine Kombination von Typen aufweisen können. Diese Strukturtypen beruhen weitgehend auf natürlicher Selektion. ⓘ
Hüllblätter
Blütenstände haben in der Regel ein modifiziertes Blattwerk, das sich vom vegetativen Teil der Pflanze unterscheidet. In der weitesten Bedeutung des Begriffs wird jedes Blatt, das mit einem Blütenstand verbunden ist, als Deckblatt bezeichnet. Ein Hüllblatt befindet sich in der Regel an dem Knoten, an dem sich der Hauptstiel des Blütenstandes bildet, der mit der Spindel der Pflanze verbunden ist, aber es können auch andere Hüllblätter im Blütenstand selbst vorhanden sein. Sie haben eine Reihe von Funktionen, unter anderem locken sie Bestäuber an und schützen die jungen Blüten. Je nach Vorhandensein oder Fehlen von Hüllblättern und ihren Merkmalen können wir folgende Arten unterscheiden:
- Ebrakteenblütenstände: Keine Hüllblätter im Blütenstand.
- Brakteenblütenstände: Die Hüllblätter des Blütenstandes sind sehr spezialisiert, manchmal auf kleine Schuppen reduziert, geteilt oder zerteilt.
- Laubblättrige Blütenstände: Die Hüllblätter sind zwar oft verkleinert, aber unspezialisiert und sehen wie die typischen Blätter der Pflanze aus, so dass statt Blütenstand meist der Begriff Blütenstiel verwendet wird. Diese Verwendung ist technisch nicht korrekt, da diese Blätter trotz ihres "normalen" Aussehens in Wirklichkeit als Hochblätter gelten, so dass der Begriff "blattartiger Blütenstand" vorzuziehen ist.
- Blattförmig-blättrige Blütenstände: Eine Zwischenform zwischen Brakteen- und Blattblütenstand. ⓘ
Wenn viele Hüllblätter vorhanden sind und sie eng mit dem Stiel verbunden sind, wie in der Familie der Asteraceae, können die Hüllblätter zusammen als Evolvente bezeichnet werden. Verfügt der Blütenstand über eine zweite Einheit von Hüllblättern, die weiter oben am Stängel sitzen, kann man sie als Evolvente bezeichnen. ⓘ

Ebrakteat-Blütenstand.

Brakteenblütenstand von Wisteria sinensis

Brakteenblütenstand.

Brakteenblütenstand von Pedicularis verticillata.

Blattförmig-blättriger Blütenstand.

Blattförmig-blättriger Blütenstand von Rhinanthus angustifolius.

Blattförmiger Blütenstand.

Blattförmiger Blütenstand von Aristolochia clematitis. ⓘ
.svg)

.svg)

.svg)

.svg)

Endblüte
Pflanzenorgane können nach zwei verschiedenen Schemata wachsen, nämlich monopodial oder racemös und sympodial oder zymös. In Blütenständen werden diese beiden unterschiedlichen Wachstumsmuster als indeterminiert bzw. determiniert bezeichnet und geben an, ob eine Endblüte gebildet wird und wo die Blüte innerhalb des Blütenstandes beginnt.
- Unbestimmter Blütenstand: Monopodiales (traubenförmiges) Wachstum. Die Endknospe wächst weiter und bildet seitliche Blüten. Eine endständige Blüte wird nie gebildet.
- Determinierter Blütenstand: Sympodialer (zymöser) Wuchs. Die Endknospe bildet eine Endblüte und stirbt dann ab. Aus den Seitenknospen wachsen dann weitere Blüten. ⓘ
Indeterminierte und determinierte Blütenstände werden manchmal auch als offene bzw. geschlossene Blütenstände bezeichnet. Die unbestimmte Form der Blüten ist von der determinierten Blüte abgeleitet. Es wird vermutet, dass unbestimmte Blüten einen gemeinsamen Mechanismus haben, der das Wachstum der Endblüte verhindert. Ausgehend von phylogenetischen Analysen ist dieser Mechanismus unabhängig voneinander mehrfach bei verschiedenen Arten aufgetreten. ⓘ
In einem unbestimmten Blütenstand gibt es keine echte Endblüte, und der Stängel hat gewöhnlich ein rudimentäres Ende. In vielen Fällen richtet sich die letzte echte Blüte, die von der Terminalknospe (Subterminalblüte) gebildet wird, auf und scheint eine Terminalblüte zu sein. Oft ist ein Überbleibsel der Endknospe weiter oben am Stängel zu sehen. ⓘ

Unbestimmter Blütenstand mit perfekter acropetaler Reifung.

Unbestimmter Blütenstand mit acropetaler Reifung und seitlichen Blütenknospen.

Unbestimmter Blütenstand mit einer subterminalen Blüte, die die terminale Blüte simuliert (Überbleibsel vorhanden) ⓘ
In determinierten Blütenständen reift die endständige Blüte in der Regel als erste (präkursive Entwicklung), während die anderen Blüten in der Regel von der Basis des Stiels aus reifen. Dieses Muster wird als acropetale Reifung bezeichnet. Wenn die Blüten von der Spitze des Stängels aus reifen, ist die Reifung basipetal, während die Reifung divergent ist, wenn die zentralen Blüten zuerst reifen. ⓘ

Determinierter Blütenstand mit acropetaler Reifung

Determinierter Blütenstand mit basipetaler Reifung

Determinierter Blütenstand mit divergenter Reifung ⓘ
Bildet die Sprossspitze eine Terminalblüte aus und verbraucht sich dadurch, spricht man von einem geschlossenen oder determinierten Blütenstand. Die einzelnen Blütenblätter folgen dabei genau der Abfolge der vorausgegangenen Blätter (Phyllotaxis). Die Terminalblüte blüht üblicherweise zuerst auf (präkursive Entfaltung), die Seiten- oder Lateralblüten unterliegen in ihrem Aufblühen (Effloration) meist einer Förderung von der Basis zur Spitze aufwärts (akropetal), oft auch von der Spitze abwärts (basipetal), seltener hin zu beiden Seiten (divergent). Durch fehlende Wachstumsstimuli oder als Hungerform kann der Blütenstand nur reduziert zur Ausbildung kommen und ganz auf die Terminalblüte beschränkt sein. ⓘ
Phyllotaxis
Wie die Blätter können auch die Blüten am Stängel nach vielen verschiedenen Mustern angeordnet sein. Siehe 'Phyllotaxis' für ausführliche Beschreibungen ⓘ
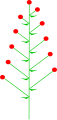
Wechselseitige Blüten

Gegenüberliegende Blüten

.svg)
Eine ähnliche Anordnung von Blättern in der Knospe wird als Ptyxis bezeichnet. ⓘ
Wenn sich eine einzelne Blüte oder ein Blütenbüschel in der Achsel eines Deckblattes befindet, wird die Lage des Deckblattes in Bezug auf den Stängel, der die Blüte(n) trägt, durch die Verwendung verschiedener Begriffe angegeben und kann ein nützlicher diagnostischer Indikator sein. ⓘ
Typische Anordnungen von Hüllblättern sind:
- Bei einigen Pflanzen befinden sich die Hüllblätter unterhalb des Blütenstandes, wo die Blüten an verzweigten Stängeln sitzen; die Hüllblätter sind nicht mit den Stängeln verbunden, die die Blüten tragen, sondern an den Hauptstängel angewachsen oder angeheftet (Angewachsen beschreibt die Verschmelzung verschiedener, nicht miteinander verbundener Teile. Wenn die miteinander verschmolzenen Teile die gleichen sind, sind sie verwachsen).
- Bei anderen Pflanzen stehen die Hüllblätter unter dem Blütenstiel oder dem Blütenstiel der Einzelblüten. ⓘ
Die metatopische Anordnung der Hüllblätter umfasst:
- Wenn das Hüllblatt am Stiel der Blüte (dem Blütenstiel) befestigt ist, nennt man es recaulescent; manchmal sind diese Hüllblätter oder Brakteolen stark verändert und scheinen Anhängsel des Blütenkelchs zu sein. Rekauleszenz ist die Verschmelzung des unterständigen Blattes mit dem Stiel, der die Knospe trägt, oder mit der Knospe selbst, so dass das Blatt oder Deckblatt mit dem Blütenstiel verwachsen ist.
- Wenn die Knospenbildung am Stängel deutlich über das Deckblatt verlagert ist, spricht man von konkazulierend. ⓘ

Blüte und Deckblatt

Lilium martagon (Blüte und Hüllblatt)

Konkauleszenz

Solanum lycopersicum (Konkauleszenz)

Rekauleszenz

Tilia cordata (Rekauleszenz) ⓘ
.svg)

.svg)

.svg)

Die den Blütenstand oder eine Einzelblüte tragende Seitenachse steht stets in der Achsel eines Tragblattes. Es kann aber auch eine Metatopie (Verlagerung) auftreten, zwei Fälle sind möglich:
- Bei der Konkauleszenz ist die Seitenachse zum Teil mit ihrer Abstammungsachse verwachsen. Dies führt dazu, dass die Blüten hier wesentlich höher am Stängel sitzen als die zugehörigen Tragblätter.
- Bei der Rekauleszenz ist die Seitenachse teilweise mit dem Stiel des Tragblattes verwachsen. Die Blüten sind in Richtung Blatt verschoben. ⓘ
Organisation
Es besteht kein allgemeiner Konsens bei der Definition der verschiedenen Blütenstände. Die folgenden Ausführungen basieren auf Focko Weberlings Morphologie der Blüten und der Blütenstände (Stuttgart, 1981). Die Hauptgruppen der Blütenstände werden nach ihrer Verzweigung unterschieden. Innerhalb dieser Gruppen sind die wichtigsten Merkmale die Achsenkreuzung und verschiedene Varianten des Modells. Sie können viele (pluriflor) oder wenige (pauciflor) Blüten enthalten. Die Blütenstände können einfach oder zusammengesetzt sein. ⓘ
Einfache Blütenstände

Unbestimmt oder traubenförmig
Unbestimmte einfache Blütenstände werden allgemein als traubenförmig bezeichnet. Der Haupttyp des traubigen Blütenstandes ist die Traube (/ˈræsiːm/, von lateinisch racemus, Traube). Die anderen Arten von traubigen Blütenständen lassen sich alle durch Erweiterung, Verdichtung, Anschwellen oder Verkleinerung der verschiedenen Achsen von diesem ableiten. Einige Übergangsformen zwischen den offensichtlichen Formen sind allgemein anerkannt.
- Eine Traube ist ein unverzweigter, unbestimmter Blütenstand mit gestielten (kurz gestielten) Blüten entlang der Achse.
- Eine Ähre ist eine Art Traube mit Blüten, die keinen Blütenstiel haben.
- Eine traubige Doldentraube ist ein unverzweigter, unbestimmter Blütenstand, der aufgrund der äußeren Blütenstiele, die immer länger sind als die inneren, eine flache oder konvexe Spitze hat.
- Eine Dolde ist eine Art von Traube mit einer kurzen Achse und mehreren gleich langen Blütenstielen, die von einem gemeinsamen Punkt auszugehen scheinen. Sie ist charakteristisch für die Doldenblütler (Umbelliferae).
- Eine Spadix ist eine Ähre mit dicht angeordneten Blüten, die von einem hochspezialisierten Hüllblatt, dem Spatel, umschlossen oder begleitet wird. Sie ist charakteristisch für die Familie der Araceae.
- Ein Blütenkopf oder Capitulum ist eine stark zusammengezogene Traube, in der die einzelnen sitzenden Blüten an einem vergrößerten Stiel sitzen. Sie ist charakteristisch für die Dipsacaceae.
- Ein Kätzchen oder Ament ist eine schuppenförmige, meist hängende Ähre oder Traube. Auch zymöse oder andere komplexe Blütenstände, die oberflächlich betrachtet ähnlich sind, werden im Allgemeinen so bezeichnet. ⓘ
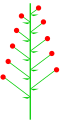
Traube

Epilobium angustifolium

Ähre

Plantago media (Ähre)

Traubenförmige Doldentraube

Iberis umbellata (traubenförmige Doldentraube)
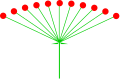
Dolde

Astrantia minor (Dolde)

Spadix

Arum maculatum (Spatel)

Kopf (rund)

Dipsacus fullonum (Kopf)

Kätzchen (traubenförmig oder stachelig)

Alnus incana (ament) ⓘ
.svg)


.svg)




.svg)

.svg)

Beim einfachen Blütenstand liegt als Verzweigungstyp ein Monopodium vor, also eine Hauptachse mit abzweigenden Nebenachsen ersten Grades (unverzweigt). Aus Tradition wird diese Verzweigungsart bei Blütenständen jedoch als razemös bezeichnet und nicht als monopodial. Der Grundtyp ist die Traube (Botrys), die anderen Blütenstände können alle durch Streckung, Stauchung, Verdickung oder Reduktion verschiedener Achsenteile aus ihr hergeleitet werden. Entsprechend häufig treten Übergangsformen auf, die zwischen den deutlich ausgebildeten Formen vermitteln. Infloreszenzen dieses Typs zählen zusammen mit den Rispentypen gemeinhin zu den namentlich bekanntesten. ⓘ
Traube
- Datei:Schirmtraube (inflorescence).svg
Schirmtraube
- Datei:Inflorescences Spike Kwiatostan Kłos.svg
Ähre, Ährchen (nur 1–3 Blüten)
- Datei:Zapfen (inflorescence).svg
Zapfen
- Datei:Kolben (inflorescence).svg
Kolben
- Datei:Koepfchen (inflorescence).svg
Köpfchen
- Datei:Inflorescences Head Kwiatostan Koszyczek.svg
Korb
- Datei:Inflorescences Umbel Kwiatostan Baldach.svg
Dolde
- Datei:Kätzchen (inflorescence).svg
Kätzchen (Trauben- oder Ährenkätzchen) ⓘ
Determiniert oder zymös
Determinierte einfache Blütenstände werden im Allgemeinen als zymös bezeichnet. Die Hauptform des zymösen Blütenstandes ist die Zyme (ausgesprochen "saim", von lateinisch cyma im Sinne von "Kohlspross", von griechisch kuma "etwas Geschwollenes"). Die Zymen werden nach diesem Schema weiter unterteilt:
- Nur eine Nebenachse: Monochasium
- Die Sekundärknospen entwickeln sich immer auf der gleichen Seite des Stängels: schraubenförmige Cyme oder Bostryx
- Die aufeinanderfolgenden Blütenstiele sind in derselben Ebene ausgerichtet: Drepanium
- Die Sekundärknospen entwickeln sich abwechselnd am Stängel: skorpioide Cyme
- Die aufeinanderfolgenden Blütenstiele sind spiralförmig angeordnet: cincinnus (charakteristisch für die Boraginaceae und Commelinaceae)
- Die aufeinanderfolgenden Blütenstiele verlaufen zickzackförmig in der gleichen Ebene: Rhipidium (viele Iridaceae)
- Die Sekundärknospen entwickeln sich immer auf der gleichen Seite des Stängels: schraubenförmige Cyme oder Bostryx
- Zwei sekundäre Achsen: dichasiale Cyme
- Sekundäre Achse noch dichasial: dichasium (charakteristisch für Caryophyllaceae)
- Nebenachse monochasisch: Doppelskorpion oder Doppelspirale
- Mehr als zwei sekundäre Achsen: Pleiochasium ⓘ
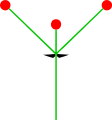
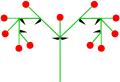
Monochasium

Doppelte Cyme
Doppelte Cyme
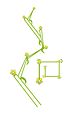
Bostryx (Seiten- und Draufsicht)

Hypericum perforatum (Bostryx)
Drepanium (Seiten- und Draufsicht)

Gladiolus imbricatus (Drepanium)

Cincinnus (Seiten- und Draufsicht)

Symphytum officinale (Zinnkraut)

Rhipidium (Seiten- und Draufsicht)

Canna sp. (Rhipidium)

Dichasium
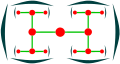
Dichasium, Draufsicht

Silene dioica (Dichasium) ⓘ
.svg)
.svg)
.svg)
.jpg)

.jpg)



.jpg)

.svg)
_(inflorescence).svg)

Eine Trugdolde kann auch so zusammengedrückt sein, dass sie wie eine Dolde aussieht. Streng genommen könnte man diese Art von Blütenstand als doldenförmige Cyme bezeichnen, obwohl sie normalerweise einfach "Dolde" genannt wird. ⓘ
Eine andere Art von definitiv einfachem Blütenstand ist der traubenförmige oder botryoide Blütenstand, d. h. eine Traube mit einer endständigen Blüte, die meist fälschlicherweise als "Traube" bezeichnet wird. ⓘ
Doldenblütiger Zyklus

Pelargonium zonale (Doldenblütiger Blütenstand)

Botryoide

Berberis vernae (botryoid) ⓘ

.svg)

Eine reduzierte Traube oder Zyme, die in der Achsel eines Hüllblattes wächst, wird als Faszikel bezeichnet. Ein Verticillaster ist ein Faszikel mit der Struktur eines Dichasiums; er ist bei den Lamiaceae weit verbreitet. Viele Verticillaster mit reduzierten Hüllblättern können einen ährenartigen Blütenstand bilden, der gemeinhin als Ähre bezeichnet wird. ⓘ

Gentiana lutea (Faszikel)

Lamium orvala (Vertikillaster)

Mentha longifolia ("Ähre") ⓘ



Zusammengesetzte Blütenstände
Einfache Blütenstände bilden die Grundlage für zusammengesetzte Blütenstände oder Synfloreszenzen. Die Einzelblüten werden dort durch einen einfachen Blütenstand ersetzt, der sowohl traubenförmig als auch zymös sein kann. Zusammengesetzte Blütenstände bestehen aus verzweigten Stängeln und können komplizierte Anordnungen aufweisen, die sich nur schwer bis zum Hauptzweig zurückverfolgen lassen. ⓘ
Eine Art zusammengesetzter Blütenstand ist der doppelte Blütenstand, bei dem sich die Grundstruktur anstelle von Einzelblüten wiederholt. Eine doppelte Traube ist beispielsweise eine Traube, in der die Einzelblüten durch andere einfache Trauben ersetzt werden; dieselbe Struktur kann wiederholt werden, um dreifache oder komplexere Strukturen zu bilden. ⓘ
Zusammengesetzte Traubenblütenstände können entweder mit einer letzten Traube enden (homöothetisch) oder nicht (heterothetisch). Eine zusammengesetzte Traube wird oft als Rispe bezeichnet. Diese Definition unterscheidet sich jedoch stark von derjenigen von Weberling. ⓘ
Zusammengesetzte Dolden sind Dolden, bei denen die Einzelblüten durch viele kleinere Dolden, die so genannten Doldentrauben, ersetzt werden. Der Stiel, der die Seitendolden mit dem Hauptstiel verbindet, wird Strahlen genannt. ⓘ

Homöotische zusammengesetzte Traube

Melilotus officinalis (homöothetische Korbblütler)
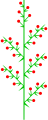
Heterothetische zusammengesetzte Traube

Hebe albicans (heterothetischer Korbblütler)

Zusammengesetzte Ähre

Lolium temulentum (zusammengesetzte Ähre)

Zusammengesetztes Körbchen (Capitulum)

Echinops ritro (zusammengesetztes Körbchen)

Zusammengesetzte (doppelte) Dolde

Laserpitium latifolium (Doppeldolde)
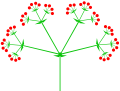
Zusammengesetzte (dreifache) Dolde ⓘ
.svg)




.svg)



.svg)
Die häufigste Art eines zusammengesetzten Blütenstandes ist die Rispe (von Webeling, oder "rispiger Blütenstand"). Eine Rispe ist ein geschlossener Blütenstand, der von oben nach unten immer stärker und unregelmäßiger verzweigt ist und in dem jede Verzweigung eine endständige Blüte trägt. ⓘ
Die so genannte cymose Doldentraube ähnelt der traubigen Doldentraube, hat aber eine rispenartige Struktur. Eine andere Art von Rispe ist die Anthela. Eine Anthela ist eine zymöse Doldentraube, bei der die seitlichen Blüten höher stehen als die zentralen. ⓘ

Rispe

Vitis vinifera (Rispe)
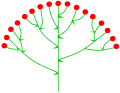
Zymöse Doldentraube

Sambucus nigra (traubige Doldentraube)

Anthela

Juncus inflexus (Ameisenblume) ⓘ


.svg)

.svg)

Eine Traube, in der die Einzelblüten durch Trugdolden ersetzt sind, wird als (unbestimmte) Thyrse bezeichnet. Die sekundären Trugdolden können zu den verschiedenen Arten der Dichasie und Monochasie gehören. Ein Botryoid, bei dem die Einzelblüten durch Trugdolden ersetzt sind, ist ein definitiver Thyrse oder Thyrsoid. Thyrsen werden oft verwirrend als Rispen bezeichnet. ⓘ

Thyrse

Aesculus hippocastanum

Thyrsoid

Syringa vulgaris ⓘ

.svg)

Andere Kombinationen sind möglich. Zum Beispiel können die Köpfe oder Dolden in einer Doldentraube oder einer Rispe angeordnet sein. ⓘ

Achillea sp. (Köpfe in einer Doldentraube)

Hedera helix (Dolden in einer Rispe) ⓘ
_-_16.JPG)

Andere
Die Familie der Asteraceae zeichnet sich durch einen hochspezialisierten Kopf aus, der technisch als Calathidum bezeichnet wird (aber gewöhnlich als Capitulum" oder Kopf" bezeichnet wird). Die Familie Poaceae hat einen eigentümlichen Blütenstand aus kleinen Ährchen, die in Rispen oder Ähren angeordnet sind, die gewöhnlich einfach und fälschlicherweise als Ähre und Rispe bezeichnet werden. Die Gattung Ficus (Moraceae) hat einen Blütenstand, der Syconium genannt wird, und die Gattung Euphorbia hat Cyathia (sing. Cyathium), die gewöhnlich in Dolden angeordnet sind. ⓘ

Matricaria chamomilla (Kalathide)

Triticum aestivum (zusammengesetzte Ähren, "Spikes")

Oryza sativa (Ähren in einer Rispe, "Rispe")

Ficus carica (Sykonium)

Euphorbia tridentata (Cyathium)

Euphorbia cyparissias (Cyathie in einer Dolde)

Coleus (falsche Ähre) ⓘ







Bei einigen Arten sind die Blütenstände auf Korbblütler oder Pseudanthien reduziert. In diesem Fall ist es schwierig, zwischen Blütenständen und Einzelblüten zu unterscheiden. ⓘ
Entwicklung und Musterung
Entwicklung
Genetische Grundlage
Gene, die für die Entwicklung der Blütenstände verantwortlich sind, wurden bei Arabidopsis eingehend untersucht. LEAFY (LFY) ist ein Gen, das die Identität des Blütenmeristems fördert und die Entwicklung der Blütenstände in Arabidopsis reguliert. Jegliche Veränderung des Zeitpunkts der LFY-Expression kann zur Bildung unterschiedlicher Blütenstände in der Pflanze führen. Zu den Genen mit ähnlicher Funktion wie LFY gehört APETALA1 (AP1). Mutationen in LFY, AP1 und ähnlichen fördernden Genen können die Umwandlung von Blüten in Sprossen verursachen. Im Gegensatz zu LEAFY unterstützen Gene wie Terminal Flower (TFL) die Aktivität eines Inhibitors, der das Wachstum von Blüten an der Blütenstandsspitze verhindert (Initiierung des Blütenprimordiums) und so die Identität des Blütenstandsmeristems aufrechterhält. Beide Arten von Genen tragen dazu bei, die Blütenentwicklung gemäß dem ABC-Modell der Blütenentwicklung zu gestalten. Kürzlich wurden oder werden Studien durchgeführt, um Homologe dieser Gene in anderen Blumenarten zu finden. ⓘ
Umwelteinflüsse
Herbivore, die sich von Blütenständen ernähren, prägen die Blütenstände, indem sie unter anderem die Lebenszeitfitness (die Anzahl der Blüten), die Samenproduktion der Blütenstände und die Pflanzendichte verringern. Ohne diese Herbivorie bilden die Blütenstände in der Regel mehr Blütenköpfe und Samen. Auch die Temperatur kann die Entwicklung der Blütenstände in unterschiedlicher Weise beeinflussen. Hohe Temperaturen können bei bestimmten Arten die ordnungsgemäße Entwicklung der Blütenknospen beeinträchtigen oder die Knospenentwicklung verzögern, während bei anderen Arten ein Temperaturanstieg die Entwicklung der Blütenstände beschleunigen kann. ⓘ
Meristeme und Architektur der Blütenstände
Der Übergang von der vegetativen zur reproduktiven Phase einer Blüte ist mit der Entwicklung eines Blütenstandsmeristems verbunden, aus dem Blütenmeristeme hervorgehen. Die Architektur der Pflanzenblütenstände hängt davon ab, welche Meristeme zu Blüten und welche zu Trieben werden. Folglich spielen Gene, die die Identität der Blütenmeristeme regulieren, eine wichtige Rolle bei der Bestimmung der Blütenstandsarchitektur, da ihre Expressionsdomäne bestimmt, wo die Blüten der Pflanze gebildet werden. ⓘ
Auf einer größeren Ebene beeinflusst die Architektur der Blütenstände die Qualität und Quantität der Nachkommenschaft aus Selbst- und Fremdbefruchtung, da die Architektur den Bestäubungserfolg beeinflussen kann. So wurde beispielsweise nachgewiesen, dass die Blütenstände von Asclepias eine Obergrenze für die Größe haben, die durch Selbstbestäubung aufgrund von Kreuzungen zwischen Blütenständen derselben Pflanze oder zwischen Blüten desselben Blütenstandes bestimmt wird. Bei Aesculus sylvatica hat sich gezeigt, dass die häufigsten Blütenstandsgrößen auch mit der höchsten Fruchtproduktion korrelieren. ⓘ
Allgemeines
Für alle Typen von Blütenständen lassen sich zusätzlich einige typenübergreifende Charakteristika finden, die beinahe in beliebiger Kombination untereinander auftreten. Sie ergänzen die Benennung der Blütenstände zusätzlich und haben keinen Einfluss auf die Typisierung. ⓘ
Beblätterung
Die Unterscheidung zwischen dem Blütenstand als generativem und dem vegetativen Teil der Pflanze geschieht oft anhand der verschiedenartigen Beblätterung:
- Fehlen die Blätter im Bereich des Blütenstandes ganz oder teilweise und sind sie als Hochblätter (Brakteen) ausgebildet und unterscheiden sich damit von der sonstigen Beblätterung, spricht man von einer brakteosen Beblätterung oder brakteosen Infloreszenz.
- Bei laubigen Tragblättern spricht man oft von einem blühenden Spross anstatt eines Blütenstands. Da diese Blätter trotz ihres laubblattartigen Äußeren auch hochblattartige Merkmale besitzen, ist frondoser Blütenstand die treffendere Bezeichnung.
- Des Weiteren existiert eine verbindende Zwischenform, der frondo-brakteose Blütenstand.
- Im Infloreszenzbereich können, so bei vielen Holzgewächsen, aber auch Blätter ohne jede Hochblattmerkmale auftreten. Es handelt sich um Kleinlaubblätter, die sich von den regulären Laubblättern durch eine gleichmäßige Reihe an Reduktion ableiten. Man spricht hierbei von fronduloser Beblätterung, den Übergang zur frondosen Infloreszenz bildet der frondo-frondulose Blütenstand. ⓘ
Reduzierte brakteose Beblätterung
- Datei:Brakteose beblätterung 2 (inflorescence).svg
Vollständige brakteose Beblätterung
- Datei:Frondobrakteose Beblätterung (inflorescence).svg
Frondo-brakteose Beblätterung
- Datei:Frondose beblätterung (inflorescence).svg
Frondose Beblätterung ⓘ
Die veraltete strenge Aufteilung in Blütenstand (brakteos) und blühenden Spross (frondos) mit den unterschiedlichen Tragblättern ist heute einer Einteilung gewichen, bei der die verschiedenen Formen von Blütenständen in einer sinnvoll weitergefassten Einteilung mit den verschiedenen Blättern als verbindendem Element definiert werden. Ein blühender Spross sollte deshalb entsprechend stets als frondoser Blütenstand bezeichnet werden. ⓘ
Terminalblüte
Für die Ausbildung der Vegetationsspitze gibt es zwei Möglichkeiten, namentlich die, ob sich eine Terminalblüte ausbildet oder nicht. Das Vorhanden- oder Nichtvorhandensein einer Terminalblüte bei den Blütenpflanzen ist für ganze Verwandtschaftskreise charakteristisch. ⓘ
Offener Blütenstand
Bildet die Sprossspitze statt einer Blüte weiterhin Hochblätter mit Knospen in ihren Achseln und endet blind in einem meist verjüngten, rudimentären Ende, so liegt ein offener Blütenstand vor. Die angelegten Blütenknospen blühen entweder allesamt auf oder sie liegen nach obenhin in immer weiterer Reduktion bis zum undeterminierten Sprossscheitel, der sogar noch zum Weiterwachsen fähig sein kann (Proliferation). Die bei Pflanzen häufige Tendenz, dass eine fehlende terminale Spitze durch die nächstgelegene ersetzt wird (Übergipfelung), kann sich auch hier zeigen: Die Blüte unterhalb der rudimentären Sprossspitze richtet sich auf und wird scheinbar zur neuen Terminalblüte. Ist ihr lateraler Ursprung noch zu erkennen, am besten durch ein noch sichtbares Rudiment, wird sie als Subterminalblüte benannt, ist der laterale Ursprung nicht einmal mehr entwicklungsgeschichtlich nachweisbar, sondern nur noch im Vergleich mit verwandten Arten, nennt man sie Pseudoterminalblüte. ⓘ
Offener Blütenstand mit vollständiger akropetaler Effloration
- Datei:Offener Blütenstand (inflorescence) m K.svg
Offener Blütenstand mit akropetaler Effloration mit Blütenknospen
- Datei:Pseudoterminalbluete (inflorescence).svg
Offener Blütenstand mit Subterminalblüte und rudimentärem Sprossscheitel ⓘ
Ansatz der Verzweigungen
Zwar ist die Art der Verzweigung ein elementares Unterscheidungsmerkmal für die verschiedenen Blütenstände, der Ansatz der Nebenachse und ihres Tragblattes an der Blütenstandsachse hingegen sind für die Typisierung der Infloreszenz nicht von Belang. Die unterschiedlichen Ansatzmöglichkeiten richten sich nach der Stellung der Blätter. ⓘ
Disperse Verzweigung
- Datei:Traube dekussiert (inflorescence).svg
Dekussierte Verzweigung ⓘ
Klassische Einteilung
Bei der klassischen Typologie der Blütenstände dient zur Unterscheidung der Hauptgruppen die Verzweigungsart. Innerhalb dieser wird anhand der Verzweigung der Achsen und vor allem deren Modifikation der Typ bestimmt. ⓘ
Zusammengesetzte Blütenstände
Razemöse Teilblütenstände
Doppelte Botryen
Ersetzt man die Blüte durch Teilblütenstände der gleichen Basisstruktur, so erhält man ein entsprechendes doppeltes Botryum (Dibotryum). So ist zum Beispiel eine Doppeltraube eine Traube, deren Blüten durch je eine Traube ersetzt wurden. Geschieht dies nur bei den seitlichen Blüten erhält man die homöothetische Form, bildet zusätzlich auch die Hauptachse noch eine Traube aus, erhält man die heterothetische Form. Die Blüten der Teilblütenstände können wiederum durch weitere Teilblütenstände ersetzt werden, es ergibt sich eine neue Verzweigungsebene. Dies geschieht aber stets nur mit der zu Grunde liegenden Struktur. Je nach Anzahl der Wiederholungen spricht man vom Dibotryum und Tribotryum, später nur noch allgemein vom Pleiobotryum. ⓘ
homöothetische Doppeltraube
- Datei:Doppeltraube 2 (inflorescence).svg
heterothetische Doppeltraube
- Datei:Inflorescences Muktispike Kwiatostan KłosZłożony.svg
Doppelähre
- Datei:Doppelkoepfchen (inflorescence).svg
Doppelkörbchen
- Datei:Inflorescences Umbel Kwiatostan BaldachZłożony.svg
Doppeldolde
- Datei:Dreifachdolde (inflorescence).svg
Dreifachdolde, ein Tribotryum ⓘ
Rispe und Verwandte
Bei der Rispe sind der gesamte Blütenstand und die Teilblütenstände immer mit einer Terminalblüte abgeschlossen. Die Teilblütenstände sind nach unten hin zunehmend stärker und unregelmäßig verzweigt. Die Seitenäste werden gemäß ihrer Blütenanzahl als Monaden (eine Blüte), Diaden (zwei Blüten) oder Triaden (drei Blüten) bezeichnet, sind sie wie eine eigenständige Rispe stark verzweigt spricht man von Spezialrispen. Insgesamt ergibt sich so eine Kegelform. Durch eine entsprechende Streckung der Seitenäste weicht dieses Erscheinungsbild einer ebenen oder leicht gewölbten Form, der Schirmrispe, und bei stärkerer Überstreckung einem trichterförmigen Aussehen bei der Spirre. Verarmt eine Rispe an Verzweigungen, so sieht sie wie eine Traube aus, nur ein eventuell verbliebener verzweigter Seitenast und vor allem die immer vorhandene Terminalblüte machen sicher deutlich, dass es sich zweifelsfrei um eine Rispe handelt. Wegen der Ähnlichkeit zur Traube (Botrys) spricht man hier vom Botryoid (Bei ährenartiger Form Stachyoid). Mit dem Verlust der Terminalblüte ist schließlich der Weg der Reduktion zur Traube vollständig beschritten. ⓘ
Rispe
- Datei:Schirmrispe (inflorescence).svg
Schirmrispe
- Datei:Spirre (inflorescence).svg
Spirre oder Trichterrispe
- Datei:Botryoid (inflorescence).svg
Botryoid, mit angedeuteter Verzweigung ⓘ
Zymöse Teilblütenstände
Beim zymösen Teilblütenstand oder kurz Zyme liegt als Verzweigungstyp ein Sympodium vor. Die Hauptachse endet mit einer Blüte, die aus den Vorblättern abzweigenden Nebenachsen übergipfeln oft die Endblüte, verzweigen weiter und enden daraufhin ebenso mit einer Blüte. Je nach Anzahl und Art der Verzweigungen, die von einer Achse entspringen, werden die verschiedenen Partialinfloreszenzen unterschieden:
- Zwei Nebenachsen, dichasial:
- Zwei transversale Vorblätter, zwei Seitenachsen: Dichasium
- Zwei transversale Vorblätter, erste Verzweigung mit zwei Seitenachsen, dann nur mehr eine Seitenachse ausgebildet: Doppelwickel oder Doppelschraubel
- Eine Nebenachse, monochasial:
- Zwei transversale Vorblätter, jedoch nur eine Seitenachse ausgebildet: Wickel oder Schraubel
- Ein medianes Vorblatt, eine Seitenachse: Fächel oder Sichel
Die Typen mit zwei Vorblättern treten bei den Zweikeimblättrigen auf, selten bei Einkeimblättrigen, ein Vorblatt umgekehrt bei Einkeimblättrigen und selten bei Zweikeimblättrigen. ⓘ
Da sich die Strukturen in der Seitenansicht nicht eindeutig voneinander unterscheiden lassen, ist zusätzlich noch der schematische Aufbau von oben dargestellt. ⓘ
Dichasium
- Datei:Dichasium (top view) (inflorescence).svg
Dichasium, schematisch von oben
- Datei:Doppelwickel (inflorescence).svg
Doppelwickel
- Datei:Doppelschraubel (inflorescence).svg
Doppelschraubel
- Datei:Wickel2 (inflorescence).svg
Wickel
- Datei:Wickel (top view) (inflorescence).svg
Wickel, schematisch von oben
- Datei:Schraubel (inflorescence).svg
Schraubel
- Datei:Schraubel (top view) (inflorescence).svg
Schraubel, schematisch von oben
- Datei:Wickel2 (inflorescence).svg
Fächel
- Datei:Fächel (top view) (inflorescence).svg
Fächel, schematisch von oben
- Datei:Schraubel (inflorescence).svg
Sichel
- Datei:Sichel (top view) (inflorescence).svg
Sichel, schematisch von oben ⓘ
Thyrsus
Bilden mehrere Zymen an einer razemösen Hauptachse den Blütenstand, so spricht man von einem Thyrsus. Die Hauptachse ist vom Typus her eine Traube, Ähre oder köpfchenartig gestaucht. Terminalblüten sind nicht immer vorhanden. ⓘ
Thyrsus mit Dichasium-Zymen
- Datei:Wickeliger zymus (inflorescence).svg
Thyrsus mit Wickel-Zymen
- Datei:Doppelwickliger zymus (inflorescence).svg
Thyrsus mit Doppelwickel-Zymen ⓘ
Werden die zymösen Teilblütenstände ihrerseits wieder durch Thyrsen ersetzt, spricht man analog zu den Spezialrispen von Spezialthyrsen und man erhält wie bei den doppelten Botryen Doppelthyrsen oder Pleiothyrsen. Analog wird hier in homöokladische oder heterokladische Form unterschieden, einfache Thyrsen sind stets homöokladisch. ⓘ
Homöokladische Thyrse mit apikaler Reduktion
- Datei:Heterokladische Thyrse (inflorescence).svg
Heterokladische Thyrse
- Datei:Disjunkt heterokladische Thyrse (inflorescence).svg
Disjunkt heterokladische Thyrse
- Datei:Pleiothyrse (inflorescence).svg
Pleiothyrse ⓘ
Cymoid
Auch wenn die Teilblütenstände zymös verzweigt sind, ist die zu Grunde liegende Struktur immer razemös. Es gibt also keine zymösen Blütenstände. Durch passende Reduktion der Struktur kann der gesamte Blütenstand allerdings rein zymös erscheinen. Man spricht dann von einem Cymoid (Zymoid). Ausgehend von den geschlossenen Thyrsusformen werden bei diesen alle Zymen bis auf die terminal gelegenen nicht ausgebildet. Entsprechend der Anzahl der verbleibenden Zymen ergeben sich monochasiale, dichasiale oder pleoichasiale Cymoide. Durch akrotone Förderung wird die Verarmung an Zymen noch verstärkt. Werden beim Pleiochasium, das bereits einen doldenartigen Charakter zeigt, die Achsen, abgesehen von den Blütenstielen, komplett reduziert, ergibt sich eine Trugdolde, die nur noch durch die als erstes erblühende Terminalblüte als solche zu erkennen ist. ⓘ
Dichasiales Cymoid
- Datei:Doppelwickel (inflorescence).svg
Doppelwickeliges Cymoid
- Datei:Doppelschraubel (inflorescence).svg
Doppelschraubeliges Cymoid
- Datei:Wickel2 (inflorescence).svg
Wickeliges oder fächeliges Cymoid
- Datei:Schraubel (inflorescence).svg
Schraubeliges oder sichelförmiges Cymoid
- Datei:Trugdolde (inflorescence).svg
Pleiochasium
- Datei:Inflorescences Umbel Kwiatostan Baldach.svg
Trugdolde ⓘ
Kuriositäten
- Puya raimondii (Bromeliengewächse): höchste Infloreszenz der Welt.
- Titanenwurz (Aronstabgewächse): Infloreszenz ist die größte Blume der Welt.
- Gattung Corypha (Palmengewächse): größte Infloreszenz der Welt mit geschätzten 10 Millionen Blüten.
- Cephalien (von griechisch Kopf) sind auffällige Blütenstandszonen, die von einigen Kakteengewächsen gebildet werden. ⓘ