Biolumineszenz


_mating.gif)
Biolumineszenz ist die Produktion und Emission von Licht durch lebende Organismen. Sie ist eine Form der Chemolumineszenz. Biolumineszenz ist bei Wirbeltieren und wirbellosen Meerestieren sowie bei einigen Pilzen, Mikroorganismen, einschließlich einiger biolumineszenter Bakterien, und terrestrischen Arthropoden wie Glühwürmchen weit verbreitet. Bei einigen Tieren ist das Licht bakteriogen, d. h. es wird von symbiotischen Bakterien wie denen der Gattung Vibrio erzeugt; bei anderen ist es autogen, d. h. es wird von den Tieren selbst erzeugt. ⓘ
Im Allgemeinen sind an der chemischen Hauptreaktion der Biolumineszenz ein lichtemittierendes Molekül und ein Enzym beteiligt, die im Allgemeinen als Luciferin bzw. Luciferase bezeichnet werden. Da es sich hierbei um Gattungsbezeichnungen handelt, werden Luciferine und Luciferasen oft nach Art oder Gruppe unterschieden, z. B. Glühwürmchen-Luciferin. In allen charakterisierten Fällen katalysiert das Enzym die Oxidation des Luciferins. ⓘ
Bei einigen Arten benötigt die Luziferase andere Kofaktoren, wie Kalzium- oder Magnesiumionen, und manchmal auch das energietragende Molekül Adenosintriphosphat (ATP). In der Evolution variieren die Luciferine nur wenig: Eines von ihnen, Coelenterazin, kommt in 11 verschiedenen Tiergruppen vor, wobei die Tiere es in einigen Fällen über die Nahrung aufnehmen. Umgekehrt variieren die Luziferasen zwischen den verschiedenen Arten stark, was darauf hindeutet, dass die Biolumineszenz im Laufe der Evolutionsgeschichte über 40 Mal aufgetreten ist. ⓘ
Sowohl Aristoteles als auch Plinius der Ältere erwähnten, dass feuchtes Holz manchmal ein Leuchten abgibt. Viele Jahrhunderte später wies Robert Boyle nach, dass sowohl bei Holz als auch bei Glühwürmchen Sauerstoff an diesem Prozess beteiligt ist. Erst im späten neunzehnten Jahrhundert wurde die Biolumineszenz richtig erforscht. Das Phänomen ist bei verschiedenen Tiergruppen weit verbreitet, vor allem in der Meeresumwelt. An Land kommt es bei Pilzen, Bakterien und einigen Gruppen wirbelloser Tiere, einschließlich Insekten, vor. ⓘ
Tiere nutzen die Biolumineszenz unter anderem zur Tarnung durch Gegenlicht, zur Nachahmung anderer Tiere, um beispielsweise Beute anzulocken, und als Signal für andere Individuen der gleichen Art, um beispielsweise Partner anzulocken. Im Labor werden auf Luziferase basierende Systeme in der Gentechnik und der biomedizinischen Forschung eingesetzt. Forscher untersuchen auch die Möglichkeit, biolumineszente Systeme für die Straßen- und Dekorationsbeleuchtung einzusetzen, und es wurde eine biolumineszente Pflanze entwickelt. ⓘ

Ein Großteil der zu Biofluoreszenz fähigen Organismen lebt aquatisch, außer bei Fischen konnte auch bei Kopffüßern, Quallen, Schnecken und Weichtiere, sowie einigen Korallen Biolumineszenz nachgewiesen werden. ⓘ
Wird das Licht nach unmittelbar vorhergehender Absorption von Licht anderer Wellenlänge erzeugt, wird es Biofluoreszenz genannt (zur Einordnung siehe Lumineszenz; beachte Unterschied zur farbigen Reflexion (Remission)). ⓘ
Geschichte
Vor der Entwicklung der Sicherheitslampe für den Einsatz in Kohlebergwerken wurden in Großbritannien und Europa getrocknete Fischhäute als schwache Lichtquelle verwendet. Mit dieser experimentellen Form der Beleuchtung wurde die Notwendigkeit vermieden, Kerzen zu verwenden, bei denen die Gefahr einer Schlagwetterexplosion bestand. Eine weitere sichere Beleuchtungsquelle in Bergwerken waren Flaschen mit Glühwürmchen. 1920 veröffentlichte der amerikanische Zoologe E. Newton Harvey eine Monografie mit dem Titel The Nature of Animal Light, in der er die frühen Arbeiten zur Biolumineszenz zusammenfasste. Harvey stellt fest, dass Aristoteles Licht erwähnt, das von totem Fisch und Fleisch erzeugt wird, und dass sowohl Aristoteles als auch Plinius der Ältere (in seiner Naturgeschichte) Licht von feuchtem Holz erwähnen. Er berichtet auch, dass Robert Boyle mit diesen Lichtquellen experimentierte und zeigte, dass sowohl sie als auch das Glühwürmchen Luft benötigen, um Licht zu erzeugen. Harvey stellt fest, dass J. Baker 1753 den Geißeltierchen Noctiluca "als ein leuchtendes Tier" identifizierte, "das mit bloßem Auge sichtbar ist", und 1854 identifizierte Johann Florian Heller (1813-1871) Pilzstränge (Hyphen) als Lichtquelle in totem Holz. ⓘ
Tuckey beschrieb in seinem posthum erschienenen Bericht über die Expedition nach Zaire von 1818, wie er die für die Lumineszenz verantwortlichen Tiere fängt. Er erwähnt Pelluciden, Krebstiere (denen er das milchige Weiß des Wassers zuschreibt) und Krebse (Krabben und Krabben). Unter dem Mikroskop beschrieb er die "leuchtende Eigenschaft" im Gehirn, die "einem höchst glänzenden Amethysten von der Größe eines großen Stecknadelkopfes" ähnelt. ⓘ
Charles Darwin bemerkte die Biolumineszenz im Meer und beschrieb sie in seinem Tagebuch:
Als wir in diesen Breitengraden in einer sehr dunklen Nacht segelten, bot das Meer ein wunderbares und sehr schönes Schauspiel. Es wehte eine frische Brise, und jeder Teil der Oberfläche, den man tagsüber als Schaum sieht, leuchtete nun in einem blassen Licht. Das Schiff trieb vor seinem Bug zwei Wogen aus flüssigem Phosphor, und in seinem Kielwasser folgte ein milchiger Zug. So weit das Auge reichte, war der Kamm jeder Welle hell, und der Himmel über dem Horizont war durch den reflektierten Glanz dieser leuchtenden Flammen nicht so völlig verdunkelt wie der Rest des Himmels. ⓘ
Darwin beobachtete auch einen leuchtenden "Quallenfisch der Gattung Dianaea" und stellte fest, dass: "Wenn die Wellen mit hellen grünen Funken schillern, so glaube ich, dass dies im Allgemeinen auf winzige Krebstiere zurückzuführen ist. Aber es besteht kein Zweifel, dass sehr viele andere pelagische Tiere, wenn sie lebendig sind, phosphoreszieren". Er vermutete, dass "ein gestörter elektrischer Zustand der Atmosphäre" wahrscheinlich dafür verantwortlich war. Daniel Pauly merkt an, dass Darwin "mit den meisten seiner Vermutungen Glück hatte, aber nicht hier", und stellt fest, dass die Biochemie zu wenig bekannt war und dass die komplexe Evolution der betroffenen Meerestiere "zu viel des Guten gewesen wäre". ⓘ

Die Biolumineszenz erregte im Kalten Krieg die Aufmerksamkeit der US-Marine, da U-Boote in manchen Gewässern ein so helles Kielwasser erzeugen können, dass sie entdeckt werden können; im Ersten Weltkrieg wurde ein deutsches U-Boot versenkt, das auf diese Weise entdeckt worden war. Die Marine war daran interessiert, vorherzusagen, wann eine solche Entdeckung möglich sein würde, und daher ihre eigenen U-Boote so zu steuern, dass sie nicht entdeckt werden. ⓘ
Zu den Anekdoten über die Navigation durch Biolumineszenz gehört auch die des Apollo-13-Astronauten Jim Lovell, der als Marinepilot den Weg zurück zu seinem Flugzeugträger USS Shangri-La gefunden hatte, als seine Navigationssysteme ausfielen. Als er seine Kabinenbeleuchtung ausschaltete, sah er das leuchtende Kielwasser des Schiffes und konnte es anfliegen und sicher landen. ⓘ
Der französische Pharmakologe Raphaël Dubois beschäftigte sich Ende des neunzehnten Jahrhunderts mit Biolumineszenz. Er untersuchte Schnellkäfer (Pyrophorus) und die Meeresmuschel Pholas dactylus. Er widerlegte die alte Vorstellung, dass Biolumineszenz von Phosphor herrührt, und wies nach, dass der Prozess mit der Oxidation einer bestimmten Verbindung, die er Luciferin nannte, durch ein Enzym zusammenhängt. Er schickte Harvey in Zucker konservierte Siphons der Molluske. Harvey interessierte sich für die Biolumineszenz, nachdem er den Südpazifik und Japan besucht und dort phosphoreszierende Organismen beobachtet hatte. Er untersuchte das Phänomen viele Jahre lang. Mit seinen Forschungen wollte er nachweisen, dass Luciferin und die Enzyme, die es zur Lichterzeugung nutzen, zwischen den Arten austauschbar sind und dass alle biolumineszenten Organismen einen gemeinsamen Vorfahren haben. Diese Hypothese erwies sich jedoch als falsch, da die verschiedenen Organismen große Unterschiede in der Zusammensetzung ihrer lichtproduzierenden Proteine aufwiesen. Er verbrachte die nächsten 30 Jahre damit, die Bestandteile zu reinigen und zu untersuchen, aber es war dem jungen japanischen Chemiker Osamu Shimomura vorbehalten, als erster kristallines Luciferin zu gewinnen. Er benutzte das Seeglühwürmchen Vargula hilgendorfii, aber es dauerte weitere zehn Jahre, bis er die Struktur der Chemikalie entdeckte und 1957 seine Arbeit Crystalline Cypridina Luciferin veröffentlichte. Shimomura, Martin Chalfie und Roger Y. Tsien erhielten 2008 den Nobelpreis für Chemie für ihre Entdeckung und Entwicklung des grün fluoreszierenden Proteins als Instrument für die biologische Forschung. ⓘ
Harvey schrieb 1957 einen detaillierten historischen Bericht über alle Formen der Lumineszenz. Ein aktualisiertes Buch über Biolumineszenz, das auch das zwanzigste und frühe einundzwanzigste Jahrhundert abdeckt, wurde kürzlich veröffentlicht. ⓘ
Entwicklung
1932 war E. N. Harvey einer der ersten, der vorschlug, wie sich die Biolumineszenz entwickelt haben könnte. In dieser frühen Arbeit schlug er vor, dass die Proto-Biolumineszenz aus Proteinen der Atmungskette entstanden sein könnte, die fluoreszierende Gruppen enthalten. Diese Hypothese wurde inzwischen widerlegt, aber sie führte zu einem beträchtlichen Interesse an den Ursprüngen des Phänomens. Heute sind die beiden vorherrschenden Hypothesen (die beide die marine Biolumineszenz betreffen) die von Howard Seliger (1993) und Rees et al. (1998) aufgestellten. ⓘ
Seligers Theorie identifiziert die Luciferase-Enzyme als Katalysator für die Entwicklung biolumineszenter Systeme. Sie besagt, dass der ursprüngliche Zweck der Luciferasen in der Verwendung von Sauerstoffasen mit gemischter Funktion bestand. Als die frühen Vorfahren vieler Arten in tiefere und dunklere Gewässer vordrangen, begünstigte die natürliche Selektion die Entwicklung einer erhöhten Augenempfindlichkeit und verbesserter visueller Signale. Wenn die Selektion eine Mutation im Oxygenase-Enzym begünstigte, das für den Abbau von Pigmentmolekülen erforderlich ist (Moleküle, die oft mit Flecken in Verbindung gebracht werden, die dazu dienen, eine Partnerin anzulocken oder einen Räuber abzulenken), könnte dies schließlich zu einer externen Lumineszenz in Geweben geführt haben. ⓘ
Rees et al. legen anhand des marinen Luciferins Coelenterazin nahe, dass die auf Luciferine wirkende Selektion durch den Druck entstanden sein könnte, ozeanische Organismen vor potenziell schädlichen reaktiven Sauerstoffspezies (z. B. H2O2 und O2- ) zu schützen. Die Funktionsverlagerung von der Antioxidation zur Biolumineszenz erfolgte wahrscheinlich, als die Stärke der Selektion auf den Antioxidationsschutz abnahm, als sich frühe Arten weiter nach unten in der Wassersäule bewegten. In größeren Tiefen ist die Exposition gegenüber ROS deutlich geringer, ebenso wie die endogene Produktion von ROS durch den Stoffwechsel. ⓘ
Obwohl Seligers Theorie anfangs populär war, wurde sie inzwischen in Frage gestellt, insbesondere aufgrund der biochemischen und genetischen Beweise, die Rees untersucht. Klar ist jedoch, dass sich die Biolumineszenz mindestens 40 Mal unabhängig voneinander entwickelt hat. Die Biolumineszenz bei Fischen begann spätestens in der Kreidezeit. Es sind etwa 1 500 Fischarten bekannt, die biolumineszierend sind; diese Fähigkeit hat sich mindestens 27 Mal unabhängig voneinander entwickelt. Bei 17 dieser Arten wurde die Fähigkeit zur Biolumineszenz durch die Aufnahme von Leuchtbakterien aus dem Wasser entwickelt, bei den anderen entstand das Licht durch chemische Synthese. Diese Fische sind in der Tiefsee erstaunlich vielfältig geworden und steuern ihr Licht mit Hilfe ihres Nervensystems, das sie nicht nur zum Anlocken von Beute oder zum Verstecken vor Räubern, sondern auch zur Kommunikation nutzen. ⓘ
Allen biolumineszierenden Organismen ist gemeinsam, dass die Reaktion zwischen einem "Luciferin" und Sauerstoff durch eine Luciferase katalysiert wird, um Licht zu erzeugen. McElroy und Seliger schlugen 1962 vor, dass sich die biolumineszente Reaktion parallel zur Photosynthese zur Entgiftung von Sauerstoff entwickelt hat. ⓘ
Thuesen, Davis et al. zeigten 2016, dass sich die Biolumineszenz unabhängig voneinander 27 Mal innerhalb von 14 Fischkladen bei Fischen mit Strahlenflossen entwickelt hat. ⓘ
Chemischer Mechanismus

Biolumineszenz ist eine Form der Chemilumineszenz, bei der Lichtenergie durch eine chemische Reaktion freigesetzt wird. An dieser Reaktion sind ein lichtemittierendes Pigment, das Luciferin, und eine Luciferase, die Enzymkomponente, beteiligt. Aufgrund der Vielfalt der Luciferin/Luciferase-Kombinationen gibt es nur sehr wenige Gemeinsamkeiten im chemischen Mechanismus. Von den derzeit untersuchten Systemen ist der einzige vereinheitlichende Mechanismus die Rolle von molekularem Sauerstoff; oft wird gleichzeitig Kohlendioxid (CO2) freigesetzt. Die Glühwürmchen-Luciferin/Luciferase-Reaktion beispielsweise erfordert Magnesium und ATP und erzeugt CO2, Adenosinmonophosphat (AMP) und Pyrophosphat (PP) als Abfallprodukte. Es können weitere Kofaktoren erforderlich sein, z. B. Kalzium (Ca2+) für das Photoprotein Aequorin oder Magnesium (Mg2+)-Ionen und ATP für die Glühwürmchen-Luciferase. Allgemein lässt sich diese Reaktion wie folgt beschreiben:
- Luciferin + O2Oxyluciferin + Lichtenergie ⓘ
![{\displaystyle {\ce {->[Text{Luciferase}][Text{andere Cofaktoren}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8326e681e6cb943a6390fdac50948116861ed3eb)

Die Qualle Aequorea victoria nutzt anstelle einer Luciferase eine andere Art von Protein, ein so genanntes Photoprotein, in diesem Fall speziell Aequorin. Wenn Kalziumionen hinzugefügt werden, erzeugt die schnelle Katalyse einen kurzen Lichtblitz, ganz im Gegensatz zu dem lang anhaltenden Leuchten, das von der Luciferase erzeugt wird. In einem zweiten, viel langsameren Schritt wird Luciferin aus der oxidierten Form (Oxyluciferin) regeneriert, so dass es sich wieder mit Aequorin verbinden kann, um einen weiteren Blitz zu erzeugen. Photoproteine sind also Enzyme, allerdings mit einer ungewöhnlichen Reaktionskinetik. Darüber hinaus wird ein Teil des blauen Lichts, das von Aequorin in Kontakt mit Kalziumionen freigesetzt wird, von einem grün fluoreszierenden Protein absorbiert, das seinerseits grünes Licht in einem Prozess freisetzt, der als resonante Energieübertragung bezeichnet wird. ⓘ
Insgesamt ist die Biolumineszenz in der Evolutionsgeschichte über 40 Mal aufgetreten. Im Laufe der Evolution haben sich die Luziferine kaum verändert: Eines von ihnen, das Coelenterazin, ist das Leuchtpigment von neun Phyla (Gruppen sehr unterschiedlicher Organismen), darunter polyzystische Radiolarien, Cercozoa (Phaeodaria), Protozoen, Kammquallen, Cnidaria (Nesseltiere) einschließlich Quallen und Korallen, Krebstiere, Weichtiere, Pfeilwürmer und Wirbeltiere (Fische mit Strahlenflossen). Nicht alle diese Organismen synthetisieren Coelenterazin: einige von ihnen erhalten es über ihre Nahrung. Umgekehrt sind die Enzyme der Luziferase sehr unterschiedlich und werden von jeder Art anders gebildet. ⓘ
Verbreitung
Biolumineszenz ist bei Tieren weit verbreitet, vor allem im offenen Meer, z. B. bei Fischen, Quallen, Kammquallen, Krebstieren und Kopffüßern, bei einigen Pilzen und Bakterien sowie bei verschiedenen wirbellosen Landtieren einschließlich Insekten. In küstennahen marinen Lebensräumen sind schätzungsweise 2,5 % der Organismen biolumineszent, während in pelagischen Lebensräumen im östlichen Pazifik etwa 76 % der wichtigsten Taxa von Tiefseetieren in der Lage sind, Licht zu erzeugen. Die meiste Lichtemission im Meer findet im blauen und grünen Lichtspektrum statt. Einige Fische mit lockeren Kiemen emittieren jedoch rotes und infrarotes Licht, und die Gattung Tomopteris strahlt gelbes Licht aus. ⓘ
Die am häufigsten anzutreffenden biolumineszenten Organismen dürften die Dinoflagellaten in den Oberflächenschichten des Meeres sein, die für die funkelnde Phosphoreszenz verantwortlich sind, die manchmal nachts in gestörtem Wasser zu sehen ist. Mindestens 18 Gattungen zeigen Leuchtkraft. Ein anderer Effekt sind die Tausende von Quadratkilometern des Ozeans, die durch das von biolumineszenten Bakterien erzeugte Licht leuchten, bekannt als Mareel- oder Milchmeereffekt. ⓘ
In fast allen Reichen der Organismen gibt es biolumineszierende Arten, jedoch nicht unter höheren Pflanzen und Landwirbeltieren. ⓘ
| Reich | primäres oder sekundäres Leuchten ⓘ |
|---|---|
| Tiere (mehrere Stämme) | primäres Leuchten (bei Wirbeltieren nur sekundäres Leuchten) |
| Pilze (wenige Arten) | primäres Leuchten |
| höhere Pflanzen | kein Leuchten |
| Einzeller (einige) | primäres Leuchten |
| Bakterien (wenige) | primäres Leuchten |
Pelagische Zone
Biolumineszenz ist in der pelagischen Zone reichlich vorhanden, wobei die höchste Konzentration in lichtlosen Tiefen und nachts im Oberflächenwasser zu finden ist. Diese Organismen nehmen an der täglichen vertikalen Wanderung aus den dunklen Tiefen an die Oberfläche teil, wodurch sich die Population der biolumineszenten Organismen über die pelagische Wassersäule verteilt. Die Ausbreitung der Biolumineszenz über verschiedene Tiefen im pelagischen Bereich wird auf den Selektionsdruck durch Raubtiere und den Mangel an Versteckmöglichkeiten im offenen Meer zurückgeführt. In Tiefen, in die das Sonnenlicht nicht vordringt, oft unterhalb von 200 m, zeigt sich die Bedeutung der Biolumineszenz darin, dass die Organismen funktionstüchtige Augen behalten, um die Biolumineszenz zu erkennen. ⓘ
Bakterielle Symbiosen
Organismen produzieren Biolumineszenz oft selbst, selten wird sie durch äußere Phänomene erzeugt. Es gibt jedoch Fälle, in denen Biolumineszenz von bakteriellen Symbionten erzeugt wird, die eine symbiotische Beziehung mit dem Wirtsorganismus eingehen. Obwohl viele Leuchtbakterien in der Meeresumwelt freilebend sind, findet man die meisten in symbiotischen Beziehungen mit Fischen, Tintenfischen, Krustentieren usw. als Wirte. Die meisten Leuchtbakterien leben im Meer, wobei die Gattungen Photobacterium und Vibrio in der Meeresumwelt dominieren. ⓘ
In der symbiotischen Beziehung profitieren die Bakterien davon, dass sie eine Nahrungsquelle und ein Refugium zum Wachsen haben. Die Wirte erhalten diese bakteriellen Symbionten entweder aus der Umwelt, durch Laich, oder das leuchtende Bakterium entwickelt sich mit seinem Wirt weiter. Koevolutionäre Wechselwirkungen werden vermutet, da die anatomischen Anpassungen der Wirtsorganismen nur für bestimmte Leuchtbakterien spezifisch geworden sind, um die ökologische Abhängigkeit der Biolumineszenz zu gewährleisten. ⓘ
Es gibt einige im Meerwasser freilebende Leuchtbakterien, die auch auf Lebensmitteln wie Fisch, Fleisch und Eiern zu finden sind. Hierzu gehören zum Beispiel Aliivibrio fischeri und Photobakterien. Aliivibrio fischeri vermehrt sich auf toten Salzwasserfischen und lässt sich leicht beobachten, wenn man einen toten, frischen Salzhering einige Zeit kühl aufbewahrt, der dann im Dunklen stellenweise leuchtet. ⓘ
Außerdem gibt es symbiotisch lebende Leuchtbakterien, die in besonderen Organen von Meerestieren vorkommen; vor allem Angler- und Laternenfische leben in Symbiose mit Leuchtbakterien. ⓘ
Es kommen häufig Symbiosen von Tieren mit Leuchtbakterien vor. Dabei werden die Bakterien von ihren Wirten mit Nahrung und Sauerstoff versorgt und leben oft in speziellen Hauttaschen oder Körperpartien. Ein Beispiel sind die Tiefseeanglerfische. ⓘ
Benthische Zone
Die Biolumineszenz von Arten in der mesopelagischen Zone ist weitgehend erforscht, aber die benthische Zone in mesopelagischen Tiefen ist noch weitgehend unbekannt. Auch die benthischen Lebensräume in Tiefen jenseits des Mesopelagials sind aus denselben Gründen nur unzureichend erforscht. Im Gegensatz zur pelagischen Zone, wo die Lichtemission im offenen Meer ungestört ist, ist das Auftreten von Biolumineszenz in der benthischen Zone seltener. Es wird darauf zurückgeführt, dass das emittierte Licht durch eine Reihe von Quellen wie den Meeresboden sowie anorganische und organische Strukturen blockiert wird. Visuelle Signale und Kommunikation, die in der pelagischen Zone vorherrschen, wie z. B. Gegenlicht, sind im benthischen Bereich möglicherweise nicht funktional oder relevant. Die Biolumineszenz von benthischen Arten in den Tiefen des Meeresbodens ist noch wenig erforscht, da es schwierig ist, Arten in diesen Tiefen zu sammeln. ⓘ
Aquatisch lebende Tiere
Fische
Bei über 180 Fischarten konnten Wissenschaftler in einer 2014 veröffentlichten Untersuchung Biofluoreszenz nachweisen. ⓘ
Zu den bioluminiszierenden Fischen zählen unter anderem:
- Anglerfische (Antennariidae), z. B. Antennarius maculatus
- Eidechsenfische (Synodontidae), z. B. Synodus dermatogenys
- Grundeln (Gobiidae), der Gattung Eviota, wie z. B. Eviota atriventris
- Seezungen, z. B. Soleichthys heterorhinos
- Plattköpfe, der Gattung Cociella, wie z. B. Cociella hutchinsi
- „falsche Muränen“, der Gattung Kaupichthys, wie z. B. Kaupichthys brachychirus
- Sandsterngucker (Dactyloscopidae), der Gattung Gillellus, wie z. B. Gillellus uranidea
- Scheinschnapper der Gattung Scolopsis, wie z. B. Scolopsis bilineata
- Seenadeln (Syngnathidae), der Gattung Corythoichthys, wie z. B. Corythoichthys haematopterus
- der Blaue Doktorfisch (Acanthurus coeruleus)
- der Buckel-Drachenkopf (Scorpaenopsis diabolus)
- der Jamaika-Stechrochen (Urobatis jamaicensis)
- der Schwellhai (Cephaloscyllium ventriosum)
- arktische Scheibenbäuche (Liparidae) aus der Gattung Liparis ⓘ

Kaupichthys brachychirus zählt zu den „ falschen Moränen“
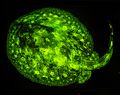
Jamaika-Stechrochen

Soleichthys heterorhinos

Blauer Doktorfisch
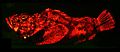
Buckel-Drachenkopf

Scheinschnapper Scolopsis bilineata

Leuchtorgane des Tiefseefisches Photostomias guernei ⓘ
.jpg)






Sonstige aquatisch lebende Tiere
Besonders verbreitet ist die Biolumineszenz unter Meeresbewohnern, vor allem in der Tiefsee (bis zu 90 Prozent der Tiefseeorganismen), aber auch in Küstengewässern (etwa fünf Prozent). Verschiedene Kopffüßer wie der Vampirtintenfisch (Vampyroteuthis infernalis), die Wunderlampen (Lycoteuthis) und andere Kalmare (Teuthida), Leuchtgarnelen (Krill, Euphausiacea), Leuchtquallen (Leuchtqualle Pelagia noctiluca, Aequorea victoria, Kronenqualle Periphylla periphylla), Vielborster (Polychaeten) wie Eusyllis blomstrandi im Helgoländer Felswatt (Helgoland), der im Sand verborgen lebende Chaetopterus variopedatus und der freischwimmende Tomopteris helgolandica, Korallen wie Renilla reniformis und verschiedene Tiefseefische. ⓘ
Unter den Nacktkiemern (Nudibranchia), meereslebenden Nacktschnecken, gibt es ebenfalls mehrere biolumineszente Arten, wie z. B. Plocamopherus imperialis und Phylliroe bucephalum. ⓘ
Insekten
Insekten mit Biolumineszenz sind zum Beispiel Leuchtkäfer (Glühwürmchen; Lampyridae) und Leuchtschnellkäfer (Gattungen Cucujo und Pyrophorus). Es gibt auch leuchtende Collembolen (Springschwänze). ⓘ
Pilze

Von über 100.000 untersuchten Pilzarten sind nur 71 biolumineszent. Dazu zählen der Honiggelbe Hallimasch (Armillaria mellea), der Leuchtende Ölbaumpilz (Omphalotus olearius) und einige Arten der Gattungen Zwergknäuelinge (Panellus, z. B. Panellus stipticus), Seitlinge (Pleurotus, z. B. Pleurotus japonicus) und Helmlinge (Mycena, z. B. Mycena citricolor, Mycena lux-coeli). ⓘ
Die Biolumineszenz entwickelte sich in vier Abstammungslinien. Es konnte gezeigt werden, dass die Biolumineszenzerscheinungen bei allen vier Abstammungslinien auf den gleichen Grundlagen beruhen. ⓘ
Einzeller

Das so genannte Meeresleuchten wird durch Plankton hervorgerufen, zum Beispiel von einzelligen Dinoflagellaten (Noctiluca scintillans), die auf Strömungsveränderungen mit der Aussendung von Licht reagieren. Meeresleuchten lässt sich an zahlreichen Küsten beobachten. ⓘ
Verwendung in der Natur
Biolumineszenz hat bei verschiedenen Taxa mehrere Funktionen. Steven Haddock et al. (2010) führen als mehr oder weniger eindeutige Funktionen bei Meeresorganismen die folgenden auf: Verteidigungsfunktionen wie Aufschrecken, Gegenlicht (Tarnung), Ablenkung (Rauchvorhang), Ablenkung von Körperteilen, Einbruchsalarm (wodurch höhere Raubtiere leichter gesehen werden können) und Warnung zur Abschreckung von Siedlern; Angriffsfunktionen wie Anlocken, Betäuben oder Verwirren von Beutetieren, Beleuchten von Beutetieren und Anlocken/Erkennen von Partnern. Für die Forscher ist es viel einfacher, festzustellen, dass eine Art in der Lage ist, Licht zu erzeugen, als die chemischen Mechanismen zu analysieren oder nachzuweisen, welche Funktion das Licht hat. In einigen Fällen ist die Funktion unbekannt, wie bei Arten aus drei Regenwurmfamilien (Oligochaeta), z. B. bei Diplocardia longa, wo die Coelomflüssigkeit Licht erzeugt, wenn sich das Tier bewegt. Die folgenden Funktionen sind bei den genannten Organismen einigermaßen gut bekannt. ⓘ
Gegenlichttarnung

Bei vielen Tieren der Tiefsee, darunter mehrere Tintenfischarten, wird die bakterielle Biolumineszenz zur Tarnung durch Gegenlicht genutzt, wobei das Tier dem von unten gesehenen Umgebungslicht angepasst wird. Bei diesen Tieren steuern Photorezeptoren die Beleuchtung so, dass sie der Helligkeit des Hintergrunds entspricht. Diese Lichtorgane sind normalerweise von dem Gewebe getrennt, das die biolumineszenten Bakterien enthält. Bei einer Art, Euprymna scolopes, sind die Bakterien jedoch ein integraler Bestandteil des Lichtorgans des Tieres. ⓘ
Anziehungskraft

Biolumineszenz wird auf vielfältige Weise und für unterschiedliche Zwecke genutzt. Der Zirren-Oktopode Stauroteuthis syrtensis nutzt Biolumineszenz aus seinen saugnapfähnlichen Strukturen. Es wird angenommen, dass sich diese Strukturen aus dem entwickelt haben, was gemeinhin als Kraken-Saugnäpfe bekannt ist. Sie haben nicht dieselbe Funktion wie die normalen Saugnäpfe, da sie aufgrund der Entwicklung von Photophoren keine Greif- oder Handhabungsfunktion mehr haben. Die Photophoren befinden sich in der Reichweite der Tiere, was Forscher zu der Vermutung veranlasst, dass sie ihre Biolumineszenz nutzen, um Beute zu fangen und anzulocken. ⓘ
Glühwürmchen nutzen Licht, um Partner anzulocken. Je nach Art gibt es zwei Systeme: Bei dem einen senden die Weibchen Licht aus ihrem Bauch aus, um Männchen anzulocken, bei dem anderen senden fliegende Männchen Signale aus, auf die die manchmal sitzenden Weibchen reagieren. Schnellkäfer senden ein orangefarbenes Licht aus dem Hinterleib aus, wenn sie fliegen, und ein grünes Licht aus dem Thorax, wenn sie gestört werden oder sich am Boden bewegen. Ersteres ist wahrscheinlich ein sexuelles Lockmittel, letzteres könnte eine Abwehrreaktion sein. Die Larven des Schnellkäfers Pyrophorus nyctophanus leben in den Oberflächenschichten von Termitenhügeln in Brasilien. Sie erhellen die Hügel durch ein helles grünliches Leuchten, das die fliegenden Insekten anlockt, von denen sie sich ernähren. ⓘ
In der Meeresumwelt ist der Einsatz von Lumineszenz zur Anlockung von Partnern vor allem bei Ostracoden bekannt, kleinen garnelenartigen Krebstieren, insbesondere aus der Familie der Cyprididae. Pheromone können für die Kommunikation über große Entfernungen eingesetzt werden, während die Biolumineszenz im Nahbereich dazu dient, die Partner anzulocken. Der Bermuda-Feuerwurm, ein polychaetischer Wurm, führt einige Nächte nach Vollmond ein kurzes Schauspiel auf, wenn das Weibchen leuchtet, um Männchen anzulocken. ⓘ
Verteidigung
_(20674653149).jpg)
Die Verteidigungsmechanismen biolumineszenter Organismen können verschiedene Formen annehmen: Aufschrecken der Beute, Gegenlicht, Rauchschleier oder Ablenkung, ablenkende Körperteile, Einbruchsalarm, Opfermarke oder Warnfärbung. Die Garnelen der Familie Oplophoridae Dana nutzen ihre Biolumineszenz, um ihre Verfolger zu erschrecken. Acanthephyra purpurea aus der Familie der Oplophoridae nutzt ihre Photophore, um Licht auszustrahlen, und kann eine biolumineszierende Substanz absondern, wenn ein Räuber in der Nähe ist. Dieser Sekretionsmechanismus ist bei Beutefischen weit verbreitet. ⓘ
Viele Kopffüßer, darunter mindestens 70 Gattungen von Tintenfischen, sind biolumineszierend. Einige Tintenfische und kleine Krustentiere verwenden biolumineszierende chemische Mischungen oder bakterielle Aufschlämmungen, so wie viele Tintenfische Tinte verwenden. Dabei wird eine Wolke aus leuchtendem Material ausgestoßen, die einen potenziellen Räuber ablenkt oder abwehrt, während das Tier in Sicherheit flieht. Der Tiefseekalmar Octopoteuthis deletron kann Teile seiner Arme, die leuchten und weiter zucken und aufblitzen, autotomisieren und so einen Räuber ablenken, während das Tier flieht. ⓘ
Dinoflagellaten können Biolumineszenz zur Verteidigung gegen Raubtiere einsetzen. Sie leuchten, wenn sie ein Raubtier entdecken, und machen so möglicherweise das Raubtier selbst angreifbarer, indem sie die Aufmerksamkeit von Raubtieren aus höheren trophischen Ebenen auf sich ziehen. Weidende Copepoden lassen alle Phytoplanktonzellen, die aufleuchten, unbeschadet wieder frei; würden sie gefressen, würden sie die Copepoden zum Leuchten bringen und Raubtiere anlocken, so dass die Biolumineszenz des Phytoplanktons der Verteidigung dient. Das Problem des leuchtenden Mageninhalts ist bei räuberischen Tiefseefischen gelöst (und die Erklärung bestätigt): Ihre Mägen haben eine schwarze Auskleidung, die verhindert, dass das Licht von biolumineszierenden Fischen, die sie verschluckt haben, größere Räuber anlockt. ⓘ
Die Seefliege ist ein kleines Krustentier, das im Sediment lebt. In Ruhe strahlt er ein dumpfes Leuchten aus, doch wenn er gestört wird, schießt er davon und hinterlässt eine Wolke aus schimmerndem blauem Licht, um Raubtiere zu verwirren. Während des Zweiten Weltkriegs wurde sie gesammelt und getrocknet, um von der japanischen Armee als Lichtquelle bei geheimen Operationen verwendet zu werden. ⓘ
Die Larven der Eisenbahnwürmer (Phrixothrix) haben an jedem Körpersegment paarige Lichtorgane, die mit grünem Licht leuchten können; man geht davon aus, dass sie der Verteidigung dienen. Sie haben auch Organe am Kopf, die rotes Licht erzeugen; sie sind die einzigen Landlebewesen, die Licht dieser Farbe aussenden. ⓘ
Warnung
Eine weit verbreitete Funktion der Biolumineszenz ist der Aposematismus, eine Warnung, dass das betreffende Lebewesen ungenießbar ist. Es wird angenommen, dass viele Glühwürmchenlarven leuchten, um Fressfeinde abzuschrecken; einige Tausendfüßler leuchten aus demselben Grund. Es wird angenommen, dass einige Meeresorganismen aus einem ähnlichen Grund Licht aussenden. Dazu gehören Schuppenwürmer, Quallen und Schlangensterne, aber es bedarf weiterer Forschung, um die Funktion der Lumineszenz vollständig zu klären. Ein solcher Mechanismus wäre für Nesseltiere mit weichem Körperbau von besonderem Vorteil, wenn sie auf diese Weise Raubtiere abschrecken könnten. Die Napfschnecke Latia neritoides ist die einzige bekannte Süßwassergastrophe, die Licht aussendet. Sie produziert grünlich leuchtenden Schleim, der möglicherweise zur Abwehr von Fressfeinden dient. Die Meeresschnecke Hinea brasiliana setzt Lichtblitze ein, wahrscheinlich um Fressfeinde abzuschrecken. Das blaugrüne Licht wird durch die durchscheinende Schale abgestrahlt, die als effizienter Lichtstreuer fungiert. ⓘ
Kommunikation

Die Kommunikation in Form von Quorum Sensing spielt bei der Regulierung der Lumineszenz bei vielen Bakterienarten eine Rolle. Kleine, extrazellulär ausgeschiedene Moleküle regen die Bakterien dazu an, Gene für die Lichterzeugung einzuschalten, wenn die Zelldichte, gemessen an der Konzentration der ausgeschiedenen Moleküle, hoch ist. ⓘ
Pyrosomen sind koloniale Tunikaten, und jeder Zooid hat ein Paar Leuchtorgane auf beiden Seiten des Einlasssiphons. Wenn diese durch Licht stimuliert werden, schalten sie sich ein und aus und verursachen ein rhythmisches Blinken. Zwischen den Zooiden verlaufen keine Nervenbahnen, aber jeder reagiert auf das von anderen Individuen erzeugte Licht und sogar auf Licht von anderen Kolonien in der Nähe. Die Kommunikation durch Lichtemission zwischen den Zooiden ermöglicht die Koordinierung der Anstrengungen der Kolonie, zum Beispiel beim Schwimmen, wo jeder Zooid einen Teil der Antriebskraft aufbringt. ⓘ
Einige bioluminöse Bakterien infizieren Nematoden, die Larven von Lepidoptera parasitieren. Wenn diese Raupen sterben, kann ihre Leuchtkraft Raubtiere zu dem toten Insekt locken und so zur Verbreitung von Bakterien und Nematoden beitragen. Ein ähnlicher Grund könnte die Ursache für die vielen Pilzarten sein, die Licht aussenden. Arten der Gattungen Armillaria, Mycena, Omphalotus, Panellus, Pleurotus und andere tun dies, indem sie meist grünliches Licht aus dem Myzel, dem Hut und den Kiemen ausstrahlen. Dies kann nachts fliegende Insekten anlocken und die Verbreitung der Sporen unterstützen, aber auch andere Funktionen können eine Rolle spielen. ⓘ
Quantula striata ist die einzige bekannte biolumineszente Landmolluske. Lichtimpulse werden von einer Drüse nahe der Vorderseite des Fußes ausgesandt und haben möglicherweise eine kommunikative Funktion, obwohl die adaptive Bedeutung nicht vollständig geklärt ist. ⓘ
Mimikry

Biolumineszenz wird von einer Vielzahl von Tieren genutzt, um andere Arten zu imitieren. Viele Arten von Tiefseefischen wie der Seeteufel und der Drachenkopf nutzen die aggressive Mimikry, um Beute anzulocken. Sie haben ein Anhängsel an ihrem Kopf, die so genannte Esca, die biolumineszente Bakterien enthält, die ein lang anhaltendes Leuchten erzeugen können, das der Fisch kontrollieren kann. Die leuchtende Esca wird baumelnd oder winkend bewegt, um kleine Tiere in die Nähe des Fisches zu locken. ⓘ
Der Zieselhai nutzt die Biolumineszenz, um seine Unterseite durch Gegenlicht zu tarnen, aber ein kleiner Bereich in der Nähe seiner Brustflossen bleibt dunkel und erscheint großen Raubfischen wie Thunfischen und Makrelen, die unter ihm schwimmen, als kleiner Fisch. Wenn sich solche Fische dem Köder nähern, werden sie von dem Hai gebissen. ⓘ
Weibliche Photuris-Glühwürmchen ahmen manchmal das Lichtmuster eines anderen Glühwürmchens, Photinus, nach, um dessen Männchen als Beute anzulocken. Auf diese Weise erhalten sie sowohl Nahrung als auch die chemischen Abwehrstoffe namens Lucibufagine, die Photuris nicht synthetisieren kann. ⓘ
Südamerikanische Riesenkakerlaken der Gattung Lucihormetica galten als das erste bekannte Beispiel für Abwehrmimikry, da sie Licht aussenden, um biolumineszente, giftige Schnellkäfer zu imitieren. Diese Behauptung ist jedoch angezweifelt worden, und es gibt keine schlüssigen Beweise dafür, dass die Schaben biolumineszent sind. ⓘ

Beleuchtung
Während die meiste marine Biolumineszenz grün bis blau ist, leuchten einige Tiefsee-Drachenköpfe der Gattungen Aristostomias, Pachystomias und Malacosteus rot. Diese Anpassung ermöglicht es den Fischen, rot pigmentierte Beutetiere zu sehen, die normalerweise für andere Organismen in der Tiefsee unsichtbar sind, da das rote Licht von der Wassersäule gefiltert wird. Der Fisch ist in der Lage, die längere Wellenlänge zu nutzen, um als Scheinwerfer für seine Beute zu fungieren, die nur er sehen kann. Es wird vermutet, dass die Fische das Licht nicht nur für die Jagd nutzen, sondern auch, um miteinander zu kommunizieren und potenzielle Partner zu finden. Die Fähigkeit der Fische, dieses Licht zu sehen, wird durch das Vorhandensein eines speziellen Rhodopsin-Pigments erklärt. Der Angler-Siphonophor (Erenna) nutzt ebenfalls rote Biolumineszenz in den Anhängseln, um Fische anzulocken. ⓘ
Der Mechanismus der Lichterzeugung erfolgt durch einen suborbitalen Photophor, der Drüsenzellen verwendet, die exergonische chemische Reaktionen erzeugen, die Licht mit einer längeren, roten Wellenlänge erzeugen. Die Drachenfischarten, die das rote Licht erzeugen, produzieren auch blaues Licht in den Photophoren auf der Rückenseite. Die Hauptfunktion dieses Pigments besteht darin, den Fisch auf die Anwesenheit seiner Beute aufmerksam zu machen. Es wird vermutet, dass das zusätzliche Pigment aus Chlorophyllderivaten der Copepoden gewonnen wird, die zu ihrer Nahrung gehören. ⓘ
Biotechnologie
Biologie und Medizin
Biolumineszierende Organismen sind ein Ziel für viele Forschungsbereiche. Luciferase-Systeme werden in der Gentechnik häufig als Reportergene eingesetzt, die durch Fluoreszenz jeweils eine andere Farbe erzeugen, und in der biomedizinischen Forschung durch Biolumineszenz-Bildgebung. So wurde beispielsweise das Glühwürmchen-Luciferase-Gen bereits 1986 für die Forschung mit transgenen Tabakpflanzen verwendet. Vibrio-Bakterien, die mit wirbellosen Meerestieren wie dem hawaiianischen Bobtail-Tintenfisch (Euprymna scolopes) eine Symbiose eingehen, sind wichtige Versuchsmodelle für Biolumineszenz. Biolumineszente aktivierte Zerstörung ist eine experimentelle Krebsbehandlung. ⓘ
Bei der In-vivo-Lumineszenz-Bildgebung von Zellen und Tieren werden Farbstoffe und fluoreszierende Proteine als Chromophore verwendet. Die Eigenschaften jedes Chromophors bestimmen, welche(r) Zellbereich(e) anvisiert und beleuchtet wird (werden). ⓘ
Lichterzeugung
Die Strukturen von Photophoren, den lichterzeugenden Organen biolumineszenter Organismen, werden von Industrieentwicklern untersucht. Die künstlich erzeugte Biolumineszenz könnte vielleicht eines Tages dazu dienen, den Bedarf an Straßenbeleuchtung zu verringern, oder zu dekorativen Zwecken, wenn es gelingt, Licht zu erzeugen, das sowohl hell genug ist als auch über lange Zeiträume zu einem vertretbaren Preis aufrechterhalten werden kann. Das Gen, das die Schwänze der Glühwürmchen zum Leuchten bringt, wurde in Senfpflanzen eingebaut. Die Pflanzen leuchten eine Stunde lang schwach, wenn man sie berührt, aber man braucht eine empfindliche Kamera, um das Leuchten zu sehen. Die Universität von Wisconsin-Madison erforscht die Verwendung gentechnisch veränderter biolumineszenter E. coli-Bakterien, die als biolumineszierende Bakterien in einer Glühbirne eingesetzt werden sollen. Im Jahr 2011 brachte Philips ein mikrobielles System für die Ambiente-Beleuchtung in Privathaushalten auf den Markt. Ein iGEM-Team aus Cambridge (England) hat damit begonnen, das Problem zu lösen, dass Luciferin bei der Lichterzeugungsreaktion verbraucht wird, indem es ein genetisches Biotechnologieteil entwickelt hat, das für ein Luciferin-regenerierendes Enzym aus dem nordamerikanischen Glühwürmchen codiert. Im Jahr 2016 begann das französische Unternehmen Glowee mit dem Verkauf von Biolumineszenz-Leuchten für Ladenfassaden und Straßenschilder, die zwischen 1 und 7 Uhr morgens eingesetzt werden können, wenn die Verwendung von Strom für diesen Zweck gesetzlich verboten ist. Sie verwendeten das biolumineszente Bakterium Aliivibrio fischeri, aber die Lebensdauer ihres Produkts betrug maximal drei Tage. Im April 2020 wurden Pflanzen gentechnisch so verändert, dass sie heller leuchten, indem Gene aus dem biolumineszenten Pilz Neonothopanus nambi verwendet wurden, um Kaffeesäure in Luciferin umzuwandeln. ⓘ
ATP-Biolumineszenz
ATP-Biolumineszenz ist der Prozess, bei dem ATP zur Erzeugung von Lumineszenz in einem Organismus verwendet wird. Sie erweist sich als ein sehr guter Biosensor zur Prüfung der Lebensfähigkeit von Zellen. Optische Biosensoren umfassen die Messung von Lumineszenz, Fluoreszenzabsorption oder Emission. Durch diese Messungen wird die quantitative Messung der ATP-Biolumineszenz nur zum Nachweis der Existenz lebender Mikroben eingesetzt. Da die Methode schnell und bequem ist, liefert sie Daten in Echtzeit. Sie ist schneller, wirtschaftlicher und einfacher zu handhaben. Optische Biosensoren erfassen das beobachtete optische Signal auf der Grundlage der Messung der an dem Phänomen beteiligten Photonen (Spiking). Sie korreliert also mit der Konzentration der mikrobiellen Population, die mit dieser Methode bestimmt wird. ⓘ
Unterscheidung zwischen lebenden und nicht lebenden Zellen
Bei der ATP-Biolumineszenz wird davon ausgegangen, dass alle lebenden Zellen im Laufe der Zeit während der chemischen Reaktion zwischen Luciferin und Luciferase zur Erzeugung von ATP die gleiche Menge ATP aufweisen. Dies geschieht, um die Lebensfähigkeit der Zelle zu messen, und ermöglicht es dem Forscher, die Menge der lebenden und toten Zellen in der Probe auf der Grundlage des Vorhandenseins oder Fehlens von ATP zu messen. Lebende Zellen, die ATP enthalten, erzeugen aufgrund der Luciferin-Luciferase-Reaktion in Gegenwart von ATP einen biolumineszenten Blitz. Tote Zellen erzeugen aufgrund der Abwesenheit von ATP keine Biolumineszenz. Die Höhe der Signalintensität ist für jede lebende Zelle in einer gesunden Probe konstant. Auf diese Weise wird die Gesamtzahl der lebenden Zellen in einer Probe bestimmt. ⓘ
Verfahren zur Messung der mikrobiellen Population
ATP, eine grundlegende Verbindung in der Luciferase-Reaktion, wird verbraucht, und im zweiten Schritt wird Oxyluciferin produziert. Das Oxyluciferin wird in einem angeregten Zustand produziert, der Licht erzeugt, wenn er in den Grundzustand zurückkehrt. Das emittierte Licht wird mit einem Luminometer gemessen. Die ATP-Konzentration ist direkt proportional zum ausgestrahlten Licht, das als Relative Light Units (RLU) gemessen wird. Die Empfängereigenschaft (Receiver Operating Characteristic, ROC) wird verwendet, um die Empfindlichkeit und Spezifität der Messungen zu berechnen. Es besteht eine direkte Korrelation zwischen der Lumineszenzintensität und der Konzentration des Standard-ATP. Es besteht ein direkter Zusammenhang zwischen der Biolumineszenz und der koloniebildenden Einheit (KBE). Die Konzentration von Standard-ATP und KBE ergibt also eine Standardkorrelation. Auf diese Weise wird ATP gemessen und die mikrobielle Population durch Biolumineszenz bestimmt. ⓘ
Es ist jedoch zu beachten, dass verschiedene Arten von mikrobiellen Populationen durch unterschiedliche ATP-Tests mit anderen Substraten und Reagenzien bestimmt werden. Renilla- und Gaussia-basierte Zellviabilitätstests verwenden das Substrat Coelenterazin. ⓘ
Erzeugung

Es werden zwei Formen der Biolumineszenz unterschieden: das primäre und das sekundäre Leuchten. Als primäres Leuchten wird es bezeichnet, wenn der Organismus die Lumineszenz selbst erzeugt. Als sekundäres Leuchten hingegen wird bezeichnet, wenn ein Organismus eine Symbiose mit anderen Lebewesen eingeht (etwa mit Leuchtbakterien), welche die Möglichkeit zum primären Leuchten besitzen. ⓘ
Luciferin / Luciferase
Eine häufig zur Biolumineszenz genutzte chemische Reaktion ist die exergone Oxidation von Luciferinen wie D-Luciferin mit molekularem Sauerstoff (O2), katalysiert durch Enzyme der Luciferasen. Dabei entstehen Dioxetane bzw. Dioxetanone, die unter Abgabe von Kohlenstoffdioxid zerfallen und die gespeicherte Energie in Form von Licht freisetzen. ⓘ
Sowohl die Luciferine als auch die Luciferasen sind art- oder gruppenspezifisch, also für jede Organismengruppe kennzeichnend. Dabei sind die Luciferasen offensichtlich im Laufe der Evolution aus anderen Enzymen, den Oxygenasen, hervorgegangen. Bei der Veränderung, meistens der Abspaltung von Teilgruppen an dem Luciferin, entsteht Energie, die als Lichtquant abgegeben wird. ⓘ
Foxfire-Biolumineszenz
Pilze nutzen die Foxfire-Biolumineszenz, wobei das Enzym Superoxiddismutase (SOD) zur Erzeugung von Biolumineszenz führt. ⓘ
Anwendung
Bereits im Mittelalter wurde in Form von Bakterienlampen die Biolumineszenz genutzt. Auch gab es Anleitungen, Buchstaben zu malen, die nachts leuchten und am Tag unsichtbar sind. Biolumineszenz ist nicht nur für die Grundlagenforschung von Interesse. Seit einiger Zeit werden verschiedene technische Anwendungen von Biolumineszenz routinemäßig eingesetzt. So wird Biolumineszenz etwa als risikoarme Markierungsmethode in der Molekularbiologie angewendet, die zusammen mit Fluoreszenz-Markierungen die Methode der radioaktiven Markierung weitgehend ersetzt hat. Auch als Nachweismethode in der Ökotoxikologie wird Biolumineszenz zum Nachweis und der Quantifizierung von Toxinen verwendet. Die Verwendung von Dinoflagellaten in der Strömungsforschung zum Nachweis von Turbulenzen wird diskutiert. Einige Forscher kündigen bereits selbstleuchtende Monitore auf Basis von Biolumineszenz an. ⓘ
1999 berichteten britische Zeitungen – und anschließend Medien in anderen Ländern – über angebliche Arbeiten an selbst leuchtenden Weihnachtsbäumen. Diese hat es allerdings nie gegeben. ⓘ
In jüngster Vergangenheit ist es den Bio-Ingenieuren des US-amerikanischen Unternehmens BioGlow gelungen, mit Hilfe von Genmanipulation und biolumineszierenden Enzymen eine autolumineszente Pflanze zu züchten. Ziel der Entwicklung war es, eine saubere, nachhaltige und bezahlbare pflanzliche Alternative für Lichtquellen zu generieren. Erzielt wird diese neue Eigenschaft der Pflanze dadurch, dass in das Chloroplastengenom der Ziertabak-Art Nicotiana alata marine Bakterien integriert wurden. Diese produzieren im Rahmen ihres induzierten Stoffwechsels auf natürlichem Weg Licht. 2020 gelang es Wissenschaftlern, Pflanzen mittels Geneditierung eigenständig und permanent heller leuchten zu lassen als bisher. Sie fügten dazu Gene eines biolumineszenten Pilzes zur Produktion von Luciferin in das Pflanzengenom ein. ⓘ