Großhirnrinde
| Zerebrale Rinde ⓘ | |
|---|---|
 Gewebeschnitt aus dem Gehirn eines erwachsenen Makaken (Macaca mulatta). Die Großhirnrinde ist die äußere Schicht, die in dunklem Violett dargestellt ist. Quelle: BrainMaps.org | |
 Golgi-gefärbte Neuronen in der Hirnrinde | |
| Einzelheiten | |
| Teil des | Großhirn |
| Bezeichner | |
| Lateinisch | Großhirnrinde (Cortex cerebri) |
| Anatomische Begriffe der Neuroanatomie [Bearbeiten auf Wikidata] | |
Die Großhirnrinde, auch Hirnmantel genannt, ist die äußere Schicht des neuronalen Gewebes des Großhirns beim Menschen und anderen Säugetieren. Die Großhirnrinde besteht zum größten Teil aus dem sechsschichtigen Neokortex und nur zu 10 % aus dem Allokortex. Sie ist durch die Längsspalte, die das Großhirn in die linke und die rechte Gehirnhälfte teilt, in zwei Kortexe unterteilt. Die beiden Hemisphären sind unterhalb der Rinde durch das Corpus callosum verbunden. Die Großhirnrinde ist der größte Ort der neuronalen Integration im zentralen Nervensystem. Sie spielt eine Schlüsselrolle bei Aufmerksamkeit, Wahrnehmung, Bewusstsein, Denken, Gedächtnis, Sprache und Bewusstsein. Die Großhirnrinde ist der Teil des Gehirns, der für die Kognition zuständig ist. ⓘ
Bei den meisten Säugetieren, mit Ausnahme von Kleinsäugern, die kleine Gehirne haben, ist die Großhirnrinde gefaltet, wodurch eine größere Oberfläche in dem begrenzten Volumen des Schädels entsteht. Abgesehen von der Minimierung des Gehirn- und Schädelvolumens ist die Faltung der Hirnrinde entscheidend für die Schaltkreise des Gehirns und seine funktionelle Organisation. Bei Säugetieren mit kleinen Gehirnen gibt es keine Faltung und die Hirnrinde ist glatt. ⓘ
Eine Falte oder ein Grat in der Hirnrinde wird als Gyrus (Plural Gyri) und eine Furche als Sulcus (Plural Sulci) bezeichnet. Diese Oberflächenfalten entstehen während der fötalen Entwicklung und reifen nach der Geburt durch den Prozess der Gyrifizierung weiter. Im menschlichen Gehirn ist der größte Teil der Großhirnrinde von außen nicht sichtbar, sondern liegt in den Sulci verborgen. Die großen Sulci und Gyri markieren die Unterteilung des Großhirns in die Hirnlappen (Lappen). Die vier großen Lappen sind der Frontal-, Parietal-, Okzipital- und Temporallappen. Weitere Lappen sind der limbische Lappen und die Inselrinde, die oft auch als Insellappen bezeichnet wird. ⓘ
In der menschlichen Großhirnrinde gibt es zwischen 14 und 16 Milliarden Neuronen. Diese sind in horizontalen Kortikalschichten und radial in Kortikalkolumnen und Minikolumnen organisiert. Die kortikalen Areale haben spezifische Funktionen, z. B. Bewegung im motorischen Kortex und Sehen im visuellen Kortex. Der visuelle Kortex befindet sich im Okzipitallappen. ⓘ
Obwohl das lateinische Wort cortex übersetzt schlicht ‚Rinde‘ bedeutet und Cortex (oder Kortex) eigentlich die gesamte Hirnrinde bezeichnet, wird Cortex in der Fachsprache auch einschränkend für die Großhirnrinde verwendet. Entsprechend bedeutet das Adjektiv cortical (oder kortikal) eigentlich „die gesamte Hirnrinde betreffend“, wird aber oft im engeren Sinne von „die Großhirnrinde betreffend“ verwendet. ⓘ
Die Großhirnrinde ist je nach Region nur 2 bis 5 Millimeter dick und ein Teil der grauen Substanz (Substantia grisea) des Großhirns. Die Nervenfasern der Neuronen der Großhirnrinde verlaufen unterhalb der Hirnrinde und sind Teil der weißen Substanz (Substantia alba) des Großhirns, die hier auch als Marklager bezeichnet wird. Großhirnrinde und Marklager bilden zusammen den Großhirnmantel (Pallium cerebri). Unterhalb der Großhirnrinde, subkortikal, befinden sich weitere Abschnitte grauer Substanz des Großhirns als subkortikale Kerngebiete (Basalganglien, Claustrum und Corpus amygdaloideum). ⓘ

Aufbau

Die Großhirnrinde ist die äußere Umhüllung der Oberflächen der Großhirnhemisphären und ist in Gyri genannte Erhebungen und Sulci genannte Furchen gefaltet. Beim menschlichen Gehirn ist sie zwischen zwei und drei oder vier Millimeter dick und macht 40 Prozent der Gehirnmasse aus. 90 Prozent der Großhirnrinde besteht aus dem sechsschichtigen Neokortex, die restlichen 10 Prozent aus dem Allokortex. In der Hirnrinde befinden sich zwischen 14 und 16 Milliarden Neuronen, die radial in kortikalen Kolumnen und Minikolumnen in den horizontal angeordneten Schichten der Hirnrinde organisiert sind. ⓘ
Der Neokortex lässt sich in verschiedene Kortexregionen unterteilen, die im Plural als Kortex bezeichnet werden und den motorischen Kortex und den visuellen Kortex umfassen. Etwa zwei Drittel der kortikalen Oberfläche sind in den Sulci vergraben, und der insuläre Kortex ist vollständig verborgen. Der Kortex ist am dicksten über einem Gyrus und am dünnsten am Boden eines Sulcus. ⓘ
Falten
Die Großhirnrinde ist so gefaltet, dass eine große Fläche des neuronalen Gewebes in die Grenzen des Neurocraniums passt. Beim Menschen hat jede hemisphärische Hirnrinde im gefalteten Zustand eine Gesamtoberfläche von etwa 0,12 Quadratmetern (1,3 sq ft). Die Faltung verläuft nach innen, weg von der Oberfläche des Gehirns, und ist auch auf der medialen Oberfläche jeder Hemisphäre innerhalb der Längsspalte vorhanden. Bei den meisten Säugetieren ist die Großhirnrinde mit den als Gyri bezeichneten Erhebungen und den als Sulci bezeichneten Vertiefungen oder Rillen gefaltet. Einige kleine Säugetiere, einschließlich einiger kleiner Nagetiere, haben glatte Hirnoberflächen ohne Gyrifikationen. ⓘ
Lappen

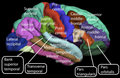
Der Cortex lässt sich grob in fünf bis sechs Lappen (Lobi) einteilen, die durch tiefere Spalten (Fissurae) voneinander getrennt sind. Hiervon liegen an der Hirnoberfläche:
- Frontallappen oder Stirnlappen (Lobus frontalis)
- Parietallappen oder Scheitellappen (Lobus parietalis)
- Occipitallappen oder Hinterhauptslappen (Lobus occipitalis)
- Temporallappen oder Schläfenlappen (Lobus temporalis) ⓘ
Bedeckt von Teilen des Frontal-, Parietal- und Temporallappens liegt seitlich der
- Insellappen (Lobus insularis) ⓘ
Zusätzlich fassen einige Autoren gewisse entwicklungsgeschichtlich ältere Teile des Cortex (z. B. Gyrus cinguli und Hippocampus) zusammen als sechsten
- Limbischen Lappen (Lobus limbicus) ⓘ
Die Untergliederung dieser Lappen ist nicht nur morphologisch, sondern auch funktionell von Bedeutung, da jedem Lappen ein spezielles primäres Verarbeitungsareal zukommt:
- Im großen Frontallappen liegen verschiedene Areale, von denen am wichtigsten die motorischen Zentren des Großhirns in und um den Gyrus praecentralis sind. In den rostralen (vorderen) Abschnitten liegt der präfrontale Cortex, der mit Handlungsplanung und -initiierung in Verbindung gebracht wird. Außerdem scheinen grundlegende Merkmale der Persönlichkeit hier lokalisiert zu sein.
- Nach hinten schließt sich der Parietallappen an, in dem das primäre sensible Zentrum liegt (Gyrus postcentralis).
- Am Pol des Occipitallappens liegt das Sehzentrum (Area striata)
- Auf der Innenseite des Temporallappens befindet sich das Hörzentrum (Area temporalis granulosa) in den sogenannten Heschlschen Querwindungen (Gyri temporales transversi)
- Die Inselrinde ist am wenigsten erforscht. Hier befindet sich unter anderem der primäre Geschmackscortex. Man vermutet hier auch das primäre Zentrum für basale Viszerosensibilität (Informationen aus den Eingeweiden).
- Im Limbischen Lappen (auch Limbisches System) werden alte Verschaltungsmuster prozessiert, deren prominenteste Vertreter Gedächtnisfunktionen und emotionale Prozesse sind. ⓘ
Die größeren Sulci und Gyri markieren die Unterteilung der Großhirnrinde in die Lappen des Gehirns. Es gibt vier Hauptlappen: den Frontallappen, den Parietallappen, den Temporallappen und den Okzipitallappen. Der insulare Kortex wird oft als Insellappen bezeichnet. Der limbische Lappen ist ein Rand des Kortex auf der medialen Seite jeder Hemisphäre und wird ebenfalls oft mit einbezogen. Außerdem werden drei Hirnlappen beschrieben: der parazentrale Lappen, der obere Parietallappen und der untere Parietallappen. ⓘ
Dicke
Bei den Säugetierarten haben größere Gehirne (in absoluten Zahlen, nicht nur im Verhältnis zur Körpergröße) tendenziell dickere Rinden. Die kleinsten Säugetiere, wie z. B. Spitzmäuse, haben eine neokortikale Dicke von etwa 0,5 mm; die Gehirne der größten Säugetiere, wie z. B. des Menschen und der Finnwale, haben eine Dicke von 2-4 mm. Es besteht ein annähernd logarithmisches Verhältnis zwischen Gehirngewicht und Kortikalisdicke. Die Magnetresonanztomographie des Gehirns (MRT) ermöglicht es, die Dicke der menschlichen Großhirnrinde zu messen und mit anderen Größen in Beziehung zu setzen. Die Dicke der verschiedenen Kortikalbereiche ist unterschiedlich, aber im Allgemeinen ist der sensorische Kortex dünner als der motorische. In einer Studie wurde ein positiver Zusammenhang zwischen der Kortikalisdicke und der Intelligenz festgestellt. In einer anderen Studie wurde festgestellt, dass der somatosensorische Kortex bei Migränepatienten dicker ist, obwohl nicht bekannt ist, ob dies die Folge oder die Ursache von Migräneanfällen ist. Eine spätere Studie mit einer größeren Patientenpopulation berichtet, dass sich die Kortikaldicke bei Migränepatienten nicht verändert hat. Eine genetische Störung der Großhirnrinde, bei der eine verminderte Faltung in bestimmten Bereichen zu einem Mikrogyrus führt, bei dem vier statt sechs Schichten vorhanden sind, wird in einigen Fällen mit Legasthenie in Verbindung gebracht. ⓘ
Schichten des Neokortex



Der Neokortex besteht aus sechs Schichten, nummeriert von I bis VI, von der äußersten Schicht I - nahe der Pia Mater - bis zur innersten Schicht VI - nahe der darunter liegenden weißen Substanz. Jede kortikale Schicht weist eine charakteristische Verteilung verschiedener Neuronen und deren Verbindungen zu anderen kortikalen und subkortikalen Regionen auf. Es gibt direkte Verbindungen zwischen verschiedenen kortikalen Bereichen und indirekte Verbindungen über den Thalamus. ⓘ
Eines der deutlichsten Beispiele für die kortikale Schichtung ist die Gennari-Linie im primären visuellen Kortex. Dabei handelt es sich um ein Band aus weißem Gewebe, das mit bloßem Auge im calcarinen Sulcus des Okzipitallappens beobachtet werden kann. Die Gennari-Linie besteht aus Axonen, die visuelle Informationen aus dem Thalamus in die Schicht IV des visuellen Kortex leiten. ⓘ
Die Anfärbung von Querschnitten der Hirnrinde, um die Lage der neuronalen Zellkörper und der intrakortikalen Axonbahnen sichtbar zu machen, ermöglichte es den Neuroanatomen zu Beginn des 20. Jahrhunderts, eine detaillierte Beschreibung der laminaren Struktur der Hirnrinde bei verschiedenen Arten zu erstellen. Korbinian Brodmann (1909) stellte fest, dass der Neokortex von Säugetieren konsequent in sechs Schichten unterteilt ist. ⓘ
Schicht I
Schicht I ist die Molekularschicht und enthält nur wenige verstreute Neuronen, darunter GABAerge Hagebuttenneuronen. Die Schicht I besteht größtenteils aus Ausläufern der apikalen dendritischen Büschel von Pyramidalneuronen und horizontal ausgerichteten Axonen sowie aus Gliazellen. Während der Entwicklung sind in dieser Schicht Cajal-Retzius-Zellen und Zellen der subpialen Körnerschicht zu finden. Auch einige stachelige Sternzellen sind hier zu finden. Es wird angenommen, dass die Eingänge zu den apikalen Büscheln entscheidend für die Rückkopplungsinteraktionen in der Großhirnrinde sind, die beim assoziativen Lernen und bei der Aufmerksamkeit eine Rolle spielen. Während man früher annahm, dass der Input für die Schicht I von der Großhirnrinde selbst stammt, weiß man heute, dass die Schicht I über den Mantel der Großhirnrinde einen erheblichen Input von Matrix- oder M-Typ-Thalamuszellen erhält (im Gegensatz zu Kern- oder C-Typ-Zellen, die zur Schicht IV gehen). ⓘ
Schicht II
Die Schicht II, die äußere Körnerschicht, enthält kleine Pyramidenneuronen und zahlreiche stellate Neuronen. ⓘ
Schicht III
Schicht III, die äußere Pyramidenschicht, enthält überwiegend kleine und mittelgroße pyramidale Neuronen sowie nicht-pyramidale Neuronen mit vertikal ausgerichteten intrakortikalen Axonen; die Schichten I bis III sind das Hauptziel der interhemisphärischen kortikalen Afferenzen, und Schicht III ist die Hauptquelle der kortikokortikalen Efferenzen. ⓘ
Schicht IV
Schicht IV, die innere Granularschicht, enthält verschiedene Typen von Stellat- und Pyramidenzellen und ist das Hauptziel thalamokortikaler Afferenzen von Neuronen des Thalamus vom Typ C (Kerntyp) sowie intrahemisphärischer kortikokortikaler Afferenzen. Die Schichten oberhalb der Schicht IV werden auch als supragranuläre Schichten (Schichten I-III) bezeichnet, während die Schichten darunter als infragranuläre Schichten (Schichten V und VI) bezeichnet werden. ⓘ
Schicht V
Schicht V, die interne Pyramidenschicht, enthält große Pyramidenneuronen. Die Axone dieser Neuronen verlassen den Kortex und verbinden sich mit subkortikalen Strukturen wie den Basalganglien. Im primären motorischen Kortex des Frontallappens enthält die Schicht V riesige Pyramidenzellen, die so genannten Betz-Zellen, deren Axone durch die innere Kapsel, den Hirnstamm und das Rückenmark wandern und den kortikospinalen Trakt bilden, der den Hauptweg für die willkürliche motorische Kontrolle darstellt. ⓘ
Schicht VI
Die Schicht VI, die polymorphe oder vielgestaltige Schicht, enthält wenige große pyramidale Neuronen und viele kleine spindelförmige pyramidale und vielgestaltige Neuronen; die Schicht VI sendet efferente Fasern zum Thalamus und stellt eine sehr präzise wechselseitige Verbindung zwischen dem Kortex und dem Thalamus her. Das heißt, Neuronen der Schicht VI aus einer kortikalen Säule verbinden sich mit Neuronen des Thalamus, die Input für dieselbe kortikale Säule liefern. Diese Verbindungen sind sowohl erregend als auch hemmend. Neuronen senden erregende Fasern zu Neuronen im Thalamus und senden auch Kollateralen zum retikulären Thalamuskern, die dieselben oder benachbarte Neuronen im Thalamus hemmen. Eine Theorie besagt, dass die Hemmungsleistung durch den cholinergen Input in die Großhirnrinde reduziert wird, was dem Hirnstamm eine einstellbare "Verstärkungskontrolle für die Weiterleitung der lemniskalischen Inputs" ermöglicht. ⓘ
Kolumnen
Die kortikalen Schichten sind nicht einfach übereinander gestapelt; es gibt charakteristische Verbindungen zwischen verschiedenen Schichten und Neuronentypen, die sich über die gesamte Dicke des Kortex erstrecken. Diese kortikalen Mikroschaltkreise werden in kortikale Kolumnen und Minikolumnen eingeteilt. Es wurde vorgeschlagen, dass die Minisäulen die grundlegenden Funktionseinheiten des Kortex sind. Vernon Mountcastle zeigte 1957, dass sich die funktionellen Eigenschaften des Kortex zwischen seitlich benachbarten Punkten abrupt ändern, in der Richtung senkrecht zur Oberfläche jedoch kontinuierlich sind. Spätere Arbeiten lieferten Beweise für das Vorhandensein funktionell unterschiedlicher kortikaler Säulen im visuellen Kortex (Hubel und Wiesel, 1959), im auditorischen Kortex, und im assoziativen Kortex. ⓘ
Kortikale Areale, denen eine Schicht IV fehlt, werden als agranulär bezeichnet. Kortikale Bereiche, die nur eine rudimentäre Schicht IV aufweisen, werden als dysgranulär bezeichnet. Die Informationsverarbeitung in den einzelnen Schichten wird durch eine unterschiedliche zeitliche Dynamik bestimmt, wobei die Schichten II/III eine langsame 2-Hz-Oszillation aufweisen, während die Schicht V eine schnelle 10-15-Hz-Oszillation besitzt. ⓘ
Arten von Kortex
Auf der Grundlage der Unterschiede in der laminaren Organisation kann die Großhirnrinde in zwei Typen eingeteilt werden: den großen Bereich des Neokortex mit sechs Zellschichten und den viel kleineren Bereich des Allokortex mit drei oder vier Schichten:
- Der Neokortex wird auch als Isokortex oder Neopallium bezeichnet und ist der Teil der reifen Großhirnrinde mit sechs verschiedenen Schichten. Beispiele für neokortikale Bereiche sind der granuläre primäre motorische Kortex und der striatale primäre visuelle Kortex. Der Neokortex hat zwei Untertypen, den echten Isokortex und den Proisokortex, der eine Übergangsregion zwischen dem Isokortex und den Regionen des Perialokortex darstellt.
- Der Allokortex ist der Teil der Großhirnrinde mit drei oder vier Schichten und hat drei Untertypen, den Paleokortex mit drei kortikalen Schichten, den Archikortex mit vier oder fünf Schichten und einen an den Allokortex angrenzenden Übergangsbereich, den Perialokortex. Beispiele für Allokortex sind der olfaktorische Kortex und der Hippocampus. ⓘ
Zwischen dem Neokortex und dem Allokortex gibt es einen Übergangsbereich, den paralimbischen Kortex, in dem die Schichten 2, 3 und 4 verschmolzen sind. Dieser Bereich umfasst den Proisokortex des Neokortex und den Perialokortex des Allokortex. Darüber hinaus kann die Großhirnrinde in vier Lappen eingeteilt werden: den Frontallappen, den Temporallappen, den Parietallappen und den Okzipitallappen, benannt nach den darüber liegenden Schädelknochen. ⓘ
Blutversorgung und Entwässerung

Die Blutversorgung der Großhirnrinde ist Teil des zerebralen Kreislaufs. Die Arterien des Gehirns liefern das Blut, das das Großhirn durchblutet. Dieses arterielle Blut transportiert Sauerstoff, Glukose und andere Nährstoffe zur Hirnrinde. Die Hirnvenen leiten das sauerstoffarme Blut und die Stoffwechselabfälle, einschließlich Kohlendioxid, zurück zum Herzen. ⓘ
Die wichtigsten Arterien, die die Hirnrinde versorgen, sind die vordere Hirnarterie, die mittlere Hirnarterie und die hintere Hirnarterie. Die vordere Großhirnarterie versorgt die vorderen Teile des Gehirns, einschließlich des größten Teils des Frontallappens. Die mittlere Zerebralarterie versorgt die Scheitellappen, die Schläfenlappen und Teile der Hinterhauptslappen. Die mittlere Zerebralarterie teilt sich in zwei Äste, die die linke und rechte Hemisphäre versorgen und sich dort weiter verzweigen. Die hintere Großhirnarterie versorgt die Hinterhauptslappen. ⓘ
Der Circulus Willis ist das wichtigste Blutsystem, das für die Blutversorgung des Großhirns und der Großhirnrinde zuständig ist. ⓘ

Entwicklung
Die pränatale Entwicklung der Großhirnrinde ist ein komplexer und fein abgestimmter Prozess, der als Kortikogenese bezeichnet wird und durch das Zusammenspiel von Genen und Umwelt beeinflusst wird. ⓘ
Neuralrohr
Die Großhirnrinde entwickelt sich aus dem vordersten Teil, der Vorderhirnregion, des Neuralrohrs. Die Neuralplatte faltet sich und schließt sich, um das Neuralrohr zu bilden. Aus dem Hohlraum im Inneren des Neuralrohrs entwickelt sich das Ventrikelsystem, und aus den Neuroepithelzellen seiner Wände entstehen die Neuronen und Glia des Nervensystems. Aus dem vordersten (vorderen oder kranialen) Teil der Neuralplatte, dem Prosencephalon, der vor Beginn der Neurulation zu sehen ist, entstehen die Gehirnhälften und später der Kortex. ⓘ
Entwicklung der kortikalen Neuronen
Die kortikalen Neuronen entstehen in der ventrikulären Zone, die sich neben den Ventrikeln befindet. Diese Zone enthält zunächst neurale Stammzellen, die in radiale Gliazellen - Vorläuferzellen - übergehen, die sich teilen, um Gliazellen und Neuronen zu bilden. ⓘ
Radiale Glia

Die Großhirnrinde besteht aus einer heterogenen Zellpopulation, aus der verschiedene Zelltypen hervorgehen. Die meisten dieser Zellen stammen aus der radialen Glia-Migration, aus der die verschiedenen Zelltypen des Neokortex hervorgehen, und es ist ein Zeitraum, der mit einer Zunahme der Neurogenese verbunden ist. In ähnlicher Weise regelt der Prozess der Neurogenese die Laminierung zur Bildung der verschiedenen Schichten des Kortex. Während dieses Prozesses kommt es zu einer zunehmenden Einschränkung des Zellschicksals, die damit beginnt, dass frühere Vorläuferzellen jeden Zelltyp im Kortex hervorbringen und spätere Vorläuferzellen nur Neuronen der oberflächlichen Schichten hervorbringen. Dieses unterschiedliche Zellschicksal führt zu einer "Inside-out"-Topographie im Kortex mit jüngeren Neuronen in den oberflächlichen Schichten und älteren Neuronen in den tieferen Schichten. Darüber hinaus werden die laminaren Neuronen in der S- oder G2-Phase gestoppt, um eine feine Unterscheidung zwischen den verschiedenen kortikalen Schichten zu ermöglichen. Die laminare Differenzierung ist erst nach der Geburt vollständig abgeschlossen, da die laminaren Neuronen während der Entwicklung noch auf extrinsische Signale und Umweltreize reagieren. ⓘ
Obwohl die Mehrzahl der Zellen, aus denen die Hirnrinde besteht, lokal von radialen Gliazellen abstammen, gibt es eine Untergruppe von Neuronen, die aus anderen Regionen einwandern. Radialglia bringt Neuronen hervor, die pyramidenförmig sind und Glutamat als Neurotransmitter verwenden. Diese wandernden Zellen steuern jedoch Neuronen bei, die sternförmig sind und GABA als Hauptneurotransmitter verwenden. Diese GABAergen Neuronen werden von Vorläuferzellen in der medialen ganglionären Eminenz (MGE) gebildet, die über die subventrikuläre Zone tangential zum Kortex wandern. Diese Wanderung von GABAergen Neuronen ist besonders wichtig, da GABA-Rezeptoren während der Entwicklung erregend sind. Diese Erregung wird in erster Linie durch den Fluss von Chloridionen durch den GABA-Rezeptor angetrieben. Bei Erwachsenen verschieben sich jedoch die Chloridkonzentrationen, was zu einem Einstrom von Chlorid führt, der die postsynaptischen Neuronen hyperpolarisiert. Die Gliafasern, die bei den ersten Teilungen der Vorläuferzellen entstehen, sind radial ausgerichtet und erstrecken sich über die gesamte Dicke der Hirnrinde von der ventrikulären Zone bis zur äußeren, pialen Oberfläche und dienen als Gerüst für die Wanderung der Neuronen aus der ventrikulären Zone nach außen. ⓘ
Bei der Geburt sind nur sehr wenige Dendriten am Zellkörper des kortikalen Neurons vorhanden, und das Axon ist noch nicht entwickelt. Im Laufe des ersten Lebensjahres nimmt die Zahl der Dendriten drastisch zu, so dass sie bis zu hunderttausend synaptische Verbindungen mit anderen Neuronen aufnehmen können. Das Axon kann sich so entwickeln, dass es sich weit vom Zellkörper entfernt. ⓘ
Asymmetrische Teilung
Die ersten Teilungen der Vorläuferzellen sind symmetrisch, wodurch sich die Gesamtzahl der Vorläuferzellen bei jedem Mitosezyklus verdoppelt. Dann beginnen einige Vorläuferzellen, sich asymmetrisch zu teilen, wobei eine postmitotische Zelle entsteht, die entlang der radialen Gliafasern wandert und die ventrikuläre Zone verlässt, und eine Vorläuferzelle, die sich bis zum Ende der Entwicklung weiter teilt und sich dann in eine Gliazelle oder eine ependymale Zelle differenziert. Mit der Verlängerung der G1-Phase der Mitose, die als selektive Zellzyklusverlängerung bezeichnet wird, wandern die neugeborenen Neuronen in die oberflächlichen Schichten des Kortex. Die wandernden Tochterzellen werden zu den Pyramidenzellen der Großhirnrinde. Der Entwicklungsprozess ist zeitlich geordnet und wird von Hunderten von Genen und epigenetischen Regulationsmechanismen gesteuert. ⓘ
Organisation der Schichten
Der schichtweise Aufbau der reifen Großhirnrinde bildet sich im Laufe der Entwicklung. Die ersten pyramidalen Neuronen, die gebildet werden, wandern aus der ventrikulären Zone und der subventrikulären Zone zusammen mit Reelin-produzierenden Cajal-Retzius-Neuronen aus der Präplatte ein. Als Nächstes teilt eine Kohorte von Neuronen, die in die Mitte der Präplatte einwandern, diese Übergangsschicht in die oberflächliche Marginalzone, die zur Schicht I des reifen Neokortex wird, und die Subplatte, die eine mittlere Schicht, die so genannte kortikale Platte, bildet. Diese Zellen werden die tiefen Schichten des reifen Kortex, die Schichten fünf und sechs, bilden. Später geborene Neuronen wandern radial in die kortikale Platte ein, vorbei an den Neuronen der tiefen Schicht, und bilden die oberen Schichten (zwei bis vier). Die Schichten des Kortex entstehen also in einer Reihenfolge von innen nach außen. Die einzige Ausnahme von dieser Inside-out-Reihenfolge der Neurogenese bildet die Schicht I der Primaten, bei denen die Neurogenese im Gegensatz zu den Nagetieren während des gesamten Zeitraums der Kortikogenese fortgesetzt wird. ⓘ
Kortikale Musterung

Die Karte der funktionellen kortikalen Areale, zu denen auch der primäre motorische und visuelle Kortex gehören, hat ihren Ursprung in einer "Protomap", die durch molekulare Signale wie den Fibroblasten-Wachstumsfaktor FGF8 früh in der Embryonalentwicklung reguliert wird. Diese Signale regulieren die Größe, Form und Position der kortikalen Bereiche auf der Oberfläche des kortikalen Primordiums, zum Teil durch die Regulierung von Gradienten der Expression von Transkriptionsfaktoren, durch einen Prozess, der als kortikale Musterung bezeichnet wird. Beispiele für solche Transkriptionsfaktoren sind die Gene EMX2 und PAX6. Beide Transkriptionsfaktoren bilden zusammen einen gegenläufigen Gradienten der Expression. Pax6 wird in hohem Maße am rostralen lateralen Pol exprimiert, während Emx2 in hohem Maße am kaudomedialen Pol exprimiert wird. Die Etablierung dieses Gradienten ist wichtig für die richtige Entwicklung. So können Mutationen in Pax6 beispielsweise dazu führen, dass die Expressionsniveaus von Emx2 über seinen normalen Expressionsbereich hinausgehen, was letztlich zu einer Ausdehnung der normalerweise vom kaudalen medialen Kortex abgeleiteten Bereiche, wie dem visuellen Kortex, führen würde. Im Gegensatz dazu können Mutationen in Emx2 dazu führen, dass sich der Bereich, in dem Pax6 exprimiert wird, ausdehnt, was zu einer Vergrößerung der frontalen und motorischen Kortexregionen führt. Daher glauben die Forscher, dass ähnliche Gradienten und Signalzentren neben dem Kortex zur regionalen Expression dieser Transkriptionsfaktoren beitragen könnten. Zwei sehr gut untersuchte Signale für die Musterbildung im Kortex sind FGF und Retinsäure. Wenn FGFs in verschiedenen Bereichen des sich entwickelnden Kortex falsch exprimiert werden, ist die kortikale Musterung gestört. Wenn Fgf8 im vorderen Pol erhöht wird, wird Emx2 herunterreguliert, und es kommt zu einer kaudalen Verschiebung der kortikalen Region. Dies führt letztlich zu einer Ausdehnung der rostralen Regionen. Fgf8 und andere FGFs spielen also eine Rolle bei der Regulierung der Expression von Emx2 und Pax6 und zeigen, wie sich die Großhirnrinde für verschiedene Funktionen spezialisieren kann. ⓘ
Die rasche Ausdehnung der kortikalen Oberfläche wird durch die Menge der Selbsterneuerung der radialen Gliazellen reguliert und wird teilweise durch FGF- und Notch-Gene gesteuert. Während der Periode der kortikalen Neurogenese und Schichtbildung beginnt bei vielen höheren Säugetieren der Prozess der Gyrifikation, der die charakteristischen Falten der Großhirnrinde erzeugt. Die Gyrifizierung wird durch ein DNA-assoziiertes Protein Trnp1 und durch FGF- und SHH-Signale reguliert. ⓘ
Entwicklung
Von allen Hirnregionen weist die Großhirnrinde die größte evolutionäre Variation auf und hat sich am jüngsten entwickelt. Im Gegensatz zu den hochkonservierten Schaltkreisen der Medulla oblongata, die kritische Funktionen wie die Regulierung der Herz- und Atemfrequenz erfüllen, sind viele Bereiche der Großhirnrinde nicht unbedingt für das Überleben notwendig. Im Laufe der Evolution der Großhirnrinde sind daher neue Funktionsbereiche entstanden und verändert worden - insbesondere Assoziationsbereiche, die keinen direkten Input von außerhalb der Rinde erhalten. ⓘ
Eine Schlüsseltheorie der kortikalen Evolution ist in der Hypothese der radialen Einheiten und der damit verbundenen Protomap-Hypothese enthalten, die erstmals von Rakic vorgeschlagen wurde. Diese Theorie besagt, dass neue kortikale Bereiche durch die Hinzufügung neuer radialer Einheiten gebildet werden, was auf der Ebene der Stammzellen geschieht. Die Protomap-Hypothese besagt, dass die zelluläre und molekulare Identität und die Merkmale der Neuronen in jedem kortikalen Bereich von kortikalen Stammzellen, den so genannten radialen Gliazellen, in einer primordialen Karte festgelegt werden. Diese Karte wird durch sekretierte Signalproteine und nachgeschaltete Transkriptionsfaktoren kontrolliert. ⓘ
Funktion

Verbindungen
Die Großhirnrinde ist mit verschiedenen subkortikalen Strukturen wie dem Thalamus und den Basalganglien verbunden und sendet über efferente Verbindungen Informationen an sie und empfängt über afferente Verbindungen Informationen von ihnen. Die meisten sensorischen Informationen werden über den Thalamus an die Großhirnrinde weitergeleitet. Geruchsinformationen gelangen jedoch über den Riechkolben zum Geruchskortex (piriformer Kortex). Braitenberg und Schüz (1998) behaupten, dass in den primären sensorischen Arealen auf der kortikalen Ebene, wo die Eingangsfasern enden, bis zu 20 % der Synapsen von extrakortikalen Afferenzen gespeist werden, dass aber in anderen Arealen und anderen Schichten der Prozentsatz wahrscheinlich viel geringer ist. ⓘ
Kortikale Bereiche
Der Cortex kann nach zwei Gesichtspunkten unterteilt werden. Zum einen aufgrund seines histologischen Feinbaus in einen sechsschichtigen Isocortex und einen drei- bis fünfschichtigen Allocortex. Innerhalb der Cortexformen lassen sich Variationen im histologischen Feinbau feststellen, nach denen die Großhirnrinde des Menschen 1909 von Korbinian Brodmann in 52 Areale unterteilt (Brodmann-Areale oder -Felder) wurde. Ein anderer Gesichtspunkt ist das stammesgeschichtliche Alter der Hirnrinde, nachdem der Cortex in einen neueren Neocortex und die älteren Archicortex und Palaeocortex unterteilt wird. ⓘ
Im Folgenden wird der histologische Aufbau des Isocortex beschrieben. Informationen zum Archicortex finden sich z. B. unter Hippocampus. ⓘ
Allgemeiner ausgedrückt besteht der Kortex in der Regel aus drei Teilen: sensorische, motorische und assoziative Areale. ⓘ
Sensorische Bereiche


Die sensorischen Areale sind die kortikalen Bereiche, die Informationen von den Sinnen empfangen und verarbeiten. Teile der Hirnrinde, die sensorische Eingaben vom Thalamus erhalten, werden als primäre sensorische Areale bezeichnet. Für den Seh-, Hör- und Tastsinn sind der primäre visuelle Kortex, der primäre auditorische Kortex und der primäre somatosensorische Kortex zuständig. Im Allgemeinen empfangen die beiden Hemisphären Informationen von der gegenüberliegenden (kontralateralen) Seite des Körpers. So empfängt beispielsweise der rechte primäre somatosensorische Kortex Informationen von den linken Gliedmaßen und der rechte visuelle Kortex Informationen aus dem linken Gesichtsfeld. Die Organisation der sensorischen Karten im Kortex spiegelt die des entsprechenden Sinnesorgans in einer so genannten topografischen Karte wider. Benachbarte Punkte in der primären Sehrinde entsprechen zum Beispiel benachbarten Punkten auf der Netzhaut. Diese topografische Karte wird als retinotopische Karte bezeichnet. In gleicher Weise gibt es eine tonotopische Karte im primären auditorischen Kortex und eine somatotopische Karte im primären sensorischen Kortex. Diese letzte topografische Karte des Körpers auf dem hinteren zentralen Gyrus wurde als eine deformierte menschliche Darstellung, der somatosensorische Homunkulus, veranschaulicht, bei der die Größe der verschiedenen Körperteile die relative Dichte ihrer Innervation widerspiegelt. Bereiche mit starker sensorischer Innervation, wie z. B. die Fingerspitzen und die Lippen, benötigen mehr Kortikalfläche, um feinere Empfindungen zu verarbeiten. ⓘ
Motorische Bereiche
Die motorischen Areale befinden sich in beiden Hemisphären des Kortex. Die motorischen Areale stehen in engem Zusammenhang mit der Steuerung willkürlicher Bewegungen, insbesondere feinteiliger Bewegungen der Hand. Die rechte Hälfte des motorischen Areals kontrolliert die linke Körperhälfte und umgekehrt. ⓘ
Zwei Bereiche des Kortex werden gemeinhin als motorisch bezeichnet:
- der primäre motorische Kortex, der die willkürlichen Bewegungen ausführt
- Ergänzende motorische Areale und prämotorischer Kortex, die willkürliche Bewegungen auswählen. ⓘ
Darüber hinaus sind motorische Funktionen beschrieben worden für:
- Posteriorer parietaler Kortex, der willkürliche Bewegungen im Raum steuert
- Dorsolateraler präfrontaler Kortex, der entscheidet, welche willkürlichen Bewegungen nach übergeordneten Anweisungen, Regeln und selbst generierten Gedanken ausgeführt werden sollen. ⓘ
Direkt unter der Großhirnrinde befinden sich miteinander verbundene subkortikale Ansammlungen grauer Substanz, die Basalganglien (oder Kerne). Die Basalganglien erhalten Input von der Substantia nigra des Mittelhirns und den motorischen Bereichen der Großhirnrinde und senden Signale an diese beiden Orte zurück. Sie sind an der motorischen Kontrolle beteiligt. Sie befinden sich seitlich des Thalamus. Die wichtigsten Bestandteile der Basalganglien sind der Nucleus caudatus, das Putamen, der Globus pallidus, die Substantia nigra, der Nucleus accumbens und der Nucleus subthalamicus. Das Putamen und der Globus pallidus werden auch als linsenförmiger Kern bezeichnet, da sie zusammen einen linsenförmigen Körper bilden. Das Putamen und der Nucleus caudatus werden nach ihrem gestreiften Erscheinungsbild auch als Corpus striatum bezeichnet. ⓘ
Assoziationsgebiete

Die Assoziationsareale sind die Teile der Großhirnrinde, die nicht zu den primären Regionen gehören. Sie sorgen für eine sinnvolle Wahrnehmungserfahrung der Welt, ermöglichen eine effektive Interaktion und unterstützen abstraktes Denken und Sprache. Der Parietal-, der Temporal- und der Okzipitallappen - alle im hinteren Teil der Hirnrinde gelegen - integrieren sensorische Informationen und im Gedächtnis gespeicherte Informationen. Der Frontallappen oder präfrontale Assoziationskomplex ist an der Planung von Handlungen und Bewegungen sowie am abstrakten Denken beteiligt. Insgesamt sind die Assoziationsareale als verteilte Netzwerke organisiert. Jedes Netzwerk verbindet Bereiche, die über weit voneinander entfernte Regionen des Kortex verteilt sind. Einzelne Netzwerke liegen nebeneinander, so dass sich eine komplexe Reihe von miteinander verwobenen Netzwerken ergibt. Die spezifische Organisation der Assoziationsnetzwerke ist umstritten, wobei es Hinweise auf Interaktionen, hierarchische Beziehungen und Wettbewerb zwischen den Netzwerken gibt. ⓘ
Beim Menschen sind die Assoziationsnetzwerke besonders wichtig für die Sprachfunktion. In der Vergangenheit ging man davon aus, dass die Sprachfähigkeit im Broca-Areal in den Bereichen des linken inferioren frontalen Gyrus, BA44 und BA45, für den Sprachausdruck und im Wernicke-Areal BA22 für die Sprachrezeption lokalisiert ist. Es hat sich jedoch gezeigt, dass die Prozesse des Sprachausdrucks und der Sprachrezeption nicht nur in den Strukturen um den Sulcus lateralis stattfinden, sondern auch im Frontallappen, in den Basalganglien, im Kleinhirn und in der Pons. ⓘ
Klinische Bedeutung

Neurodegenerative Erkrankungen wie die Alzheimer-Krankheit und die Lafora-Krankheit zeigen als Marker eine Atrophie der grauen Substanz der Großhirnrinde. ⓘ
Andere Erkrankungen des Zentralnervensystems umfassen neurologische Störungen wie Epilepsie, Bewegungsstörungen und Sprachstörungen (Aphasie). ⓘ
Bei Hirnschäden durch Krankheiten oder Traumata kann ein bestimmter Hirnlappen geschädigt sein, wie z. B. bei einer Frontallappenstörung, und die damit verbundenen Funktionen werden beeinträchtigt. Die Blut-Hirn-Schranke, die das Gehirn vor Infektionen schützt, kann geschädigt werden, so dass Krankheitserreger eindringen können. ⓘ
Der sich entwickelnde Fötus ist anfällig für eine Reihe von Umweltfaktoren, die Geburtsfehler und Probleme in der späteren Entwicklung verursachen können. Mütterlicher Alkoholkonsum kann zum Beispiel eine fetale Alkoholspektrumstörung verursachen. Andere Faktoren, die zu Störungen der Neuroentwicklung führen können, sind Giftstoffe wie Medikamente und Strahlenbelastung durch Röntgenstrahlen. Auch Infektionen können die Entwicklung der Hirnrinde beeinträchtigen. Eine Virusinfektion ist eine der Ursachen der Lissenzephalie, die zu einer glatten Hirnrinde ohne Gyrifikation führt. ⓘ
Eine Art der Elektrokortikographie, die so genannte kortikale Stimulationskartierung, ist ein invasives Verfahren, bei dem Elektroden direkt auf das freiliegende Gehirn gelegt werden, um die Funktionen bestimmter Bereiche der Hirnrinde zu lokalisieren. Es wird in klinischen und therapeutischen Anwendungen eingesetzt, einschließlich der Kartierung vor chirurgischen Eingriffen. ⓘ
Gene im Zusammenhang mit kortikalen Störungen
Es gibt eine Reihe von Genmutationen, die eine Vielzahl von genetischen Störungen der Großhirnrinde verursachen können, darunter Mikrozephalie, Schizencephalie und verschiedene Formen der Lissencephalie. Chromosomenanomalien können auch zu einer Reihe von neurologischen Entwicklungsstörungen wie dem fragilen X-Syndrom und dem Rett-Syndrom führen. ⓘ
MCPH1 kodiert für Mikrozephalin, und Störungen in diesem Gen sowie in ASPM werden mit Mikrozephalie in Verbindung gebracht. Mutationen in dem Gen NBS1, das für Nibrin kodiert, können das Nijmegen-Breakage-Syndrom verursachen, das durch Mikrozephalie gekennzeichnet ist. ⓘ
Mutationen in EMX2 und COL4A1 werden mit Schizencephalie in Verbindung gebracht, einem Zustand, der durch das Fehlen großer Teile der Großhirnhälften gekennzeichnet ist. ⓘ
Geschichte
1909 unterschied Korbinian Brodmann verschiedene Bereiche des Neokortex auf der Grundlage zytoarchitektonischer Unterschiede und teilte die Großhirnrinde in 52 Regionen ein. ⓘ
Rafael Lorente de Nó, ein Schüler von Santiago Ramon y Cajal, identifizierte mehr als 40 verschiedene Typen von kortikalen Neuronen anhand der Verteilung ihrer Dendriten und Axone. ⓘ
Andere Tiere
Die Großhirnrinde leitet sich vom Pallium ab, einer geschichteten Struktur im Vorderhirn aller Wirbeltiere. Die Grundform des Palliums ist eine zylindrische Schicht, die mit Flüssigkeit gefüllte Ventrikel umschließt. Um den Umfang des Zylinders herum befinden sich vier Zonen, das dorsale Pallium, das mediale Pallium, das ventrale Pallium und das laterale Pallium, von denen man annimmt, dass sie mit dem Neokortex, dem Hippocampus, der Amygdala bzw. dem olfaktorischen Kortex vergleichbar sind. ⓘ
Bis vor kurzem war kein Gegenstück zur Großhirnrinde bei wirbellosen Tieren bekannt. Eine 2010 in der Fachzeitschrift Cell veröffentlichte Studie, die sich auf Genexpressionsprofile stützte, wies jedoch auf eine starke Verwandtschaft zwischen der Großhirnrinde und den Pilzkörpern des Regenwurms Platynereis dumerilii hin. Pilzkörper sind Strukturen im Gehirn vieler Arten von Würmern und Gliederfüßern, von denen bekannt ist, dass sie eine wichtige Rolle beim Lernen und beim Gedächtnis spielen. Die genetischen Beweise deuten auf einen gemeinsamen evolutionären Ursprung hin und deuten somit darauf hin, dass die Ursprünge der frühesten Vorläufer der Großhirnrinde in die präkambrische Ära zurückreichen. ⓘ
Zusätzliche Bilder
Seitliche Oberfläche der menschlichen Großhirnrinde

Mediale Oberfläche der menschlichen Großhirnrinde ⓘ


Gliederung
Makroskopische Gliederung
Die noch heute gültige Terminologie der Hirnlappen und -windungen wurde 1869 von Alexander Ecker (1816–1887) vorgeschlagen. ⓘ
Faltung (Gyrierung)

Dreieck (▼): Gehirnvolumen
Kreis (⚫): Hirnoberfläche
Raute (◆) im kleineren Schaubild: Verhältnis von Oberfläche zu Volumen ⓘ
Die Großhirnrinde zeichnet sich bei vielen Säugetieren durch zahlreiche Windungen (lateinisch-griechisch Gyri, Singular Gyrus), Spalten (lateinisch Fissurae, Singular Fissura) und Furchen (lateinisch Sulci, Singular Sulcus) aus. Die Faltung dient der Vergrößerung der Oberfläche: beim Menschen beträgt diese etwa 1800 cm². Bei der Furchung des Cortex unterscheidet man eine Primärfurchung, die bei allen Individuen annähernd gleich ist, von einer Sekundär- und Tertiärfurchung, die so individuell wie ein Fingerabdruck sein können. ⓘ
Gefurchte Gehirne bezeichnet man als gyrenzephal. Bei einigen Kleinsäugern (beispielsweise Nagetiere, Igel) und bei Vögeln besitzt der Cortex keine Furchen (lissenzephales Gehirn). Das Gen Trnp1 besitzt die Fähigkeit durch unterschiedliche Expressions-Levels die Gyrierung zu beeinflussen und in normalerweise lissenzephalen Gehirnen sogar zu induzieren. ⓘ
Die Lobi und Gyri werden durch die Fissuren und Sulci voneinander getrennt. Deren wichtigste Vertreter sind:
- Fissura longitudinalis, die den Spalt zwischen den beiden Hemisphären bildet. In die Fissura longitudinalis ragt die Falx cerebri.
- Sulcus centralis trennt den Frontal- und Parietallappen (Gyrus praecentralis beziehungsweise Gyrus postcentralis) und somit das primär motorische vom primär sensiblen Rindenfeld
- Sulcus lateralis (auch Sylvische Fissur) liegt oberhalb der Insula und trennt den Temporallappen von den darüber liegenden Frontal- und Parietallappen
- Sulcus parietooccipitalis zwischen Parietal- und Occipitallappen
- Sulcus calcarinus teilt innerhalb des Occipitallappens die primäre Sehrinde in einen oberen und unteren Anteil, welcher das jeweils gegenüberliegende Gesichtsfeld repräsentiert, das heißt oberhalb des Sulcus calcarinus das untere Gesichtsfeld, unterhalb des Sulcus calcarinus das obere. ⓘ
Funktionelle Gliederung
Konnektom
Das Konnektom gliedert funktionell nach jeweils verbundenen Neuronen, lokale Strukturen sind nebensächlich. Bei den lokalen Gliederungen bestehen die funktionellen Teile aus Nervengewebe, also aus vermengten Neuronen und Gliazellen. Das Konnektom beschreibt bisher alleinig die verbundenen Neuronen. Die Astrozyten befinden sich bei dieser Gliederung zwischen den verbundenen Neuronen. ⓘ
Verschaltung
Forschung
Das Blue-Brain-Projekt hatte eine Computersimulation des Neocortex zum Ziel. ⓘ
Die Evolution und Funktion des Großhirns
Das menschliche Gehirn ist keine Neuentwicklung der Natur. Es hat sich wie alle anderen Organe aus einfachen Formen entwickelt. Das Nervensystem entwickelt sich aus einer sehr einfachen Struktur, dem äußeren Keimblatt (Ektoderm). Dass ein Organ der Informationsverarbeitung aus der äußeren Grenzschicht entsteht, ist leicht verständlich, weil hier die Reize aus der Umwelt auftreffen. Erst im Lauf der Evolution wurden die empfindlichen Nervenverbände in die Tiefe des Neuralrohrs verlegt, weil sie dort besser geschützt sind. Die Verbindungen zur Außenwelt blieben über die nun spezialisierten Sinnesorgane bestehen. ⓘ
Mit der Entstehung spezialisierter Sinnesorgane ist die Bildung einer Nervenzentrale verbunden, die den ganzen Körper einheitlich nach den Sinneseindrücken steuern kann. Weil sich schon früh in der Geschichte der Wirbeltiere Augen, Ohren und chemische Sinne (Geschmack, Geruch) ausbilden, ist das Gehirn aller Wirbeltiere in gleicher Art zur zentralen Integration dieser Sinne konstruiert. ⓘ
Das Endhirn war zunächst Verarbeitungszentrum für das Geruchsorgan. Weil der Geruchssinn ein allgemeines Warn- und Reizsystem hoher Empfindlichkeit ist, aber wenig über die räumliche Situation bzw. den Ort der Reizquelle aussagt, ist für das Riechhirn eine Verbindung mit den optischen und akustischen Zentren des Mittelhirns notwendig, mit der alle Sinnesqualitäten auf einer gemeinsamen Ebene vereinigt werden. ⓘ
Diese gemeinsame Ebene entsteht schon bei den Reptilien aus einer Erweiterung des Endhirns als Telencephalon oder rudimentärer Cortex. Bereits bei Fröschen und Salamandern ist diese Hirnstruktur für die Integration der verschiedenartigen Reize angelegt. Für die Umschaltung der Seh-, Tast- und Hörwelt vom Mittelhirn auf das Endhirn entwickelt sich ein Teil des Vorderhirns, das Zwischenhirn. Aus ihm entsteht der Thalamus, der aus mehreren Kerngruppen die spezifischen Signale des Mittelhirns zu spezifischen Regionen der Großhirnrinde sendet. Man bezeichnet diese Anordnung als ein Projektionssystem, die Anatomen nannten den Thalamus das „Tor zum Bewusstsein“. ⓘ
Mit dem Wegfall des Schuppenkleides der Fische bzw. der Hornschuppen der Reptilien wurde bei den Säugetieren die ganze Haut zu einem empfindlichen Sinnesorgan, das ebenso über Projektionsbahnen im Cortex mit den übrigen Sinnesqualitäten in ganzheitliche Verbindung gelangt. ⓘ
Eine Nervenzentrale, in der alle Qualitäten der Umweltsignale zusammengeführt werden, wäre nicht sinnvoll, wenn in ihr keine Befehle für die Reaktionen des Organismus gebildet und an die ausführenden Organe geleitet werden könnten. Weil das Geruchsorgan von Anfang an einen steuernden Zugriff auf komplexe Verhaltensweisen hat, kann das zum Integrationszentrum aller Sinne erweiterte Riechhirn auf diese Steuerungsbahnen zurückgreifen, um aus der Vereinheitlichung aller Empfindungen ganzheitliche Verhaltensschritte zu entwickeln. ⓘ
Diese Integrationsleistung des Neocortex, die alle Sinne zu einem Ganzen verbindet und sinnvolle Verhaltensmuster daraus herstellt, ermöglicht bereits Ratten, Katzen usw. ein intelligentes Verhalten, das bei Insekten oder einfachen Organismen so nicht vorkommt. Dabei zeigt sich, dass schon Vögel und Mäuse ihr integratives Zentrum, die Hirnrinde, nicht nur als Kommandozentrale, sondern auch als besonders leistungsfähigen Informationsspeicher (Gedächtnis) nutzen können (siehe auch: Gehirn und Kognition der Vögel). Eine Fliege lernt es nie, den Zusammenstoß mit einer Fensterscheibe zu vermeiden, während ein Vogel nach einigen Erfahrungen einen vorsichtigen Umgang mit der durchsichtigen Wand lernt. ⓘ
Nur Tiere, die über einen Cortex verfügen, können auch dressiert werden, das heißt, sie entwickeln ein Gedächtnis für sprachliche Anweisungen, die auch über die angeborenen Verhaltensmuster dominieren können. Deutlich ist diese Lernfähigkeit bei den Delphinen, die als Säugetiere mit einem mächtigen Cortex ausgestattet und gut dressierbar sind, während die relativ großhirnarmen Haie zur Dressur wenig geeignet sind. ⓘ
Mit der Entwicklung des Cortex kommt zunehmend eine spielerische Phase der Jungtiere zum Vorschein, die als Lernphase der Hirnrinde zu verstehen ist und uns den Eindruck vermittelt, dass diese Tiere (z. B. Hunde, Katzen usw.) ähnliche geistige Zustände wie die Menschen empfinden. ⓘ
Eine mächtige Entwicklung der Großhirnrinde wurde bei den Affen durch die Sonderstellung der Hände ausgelöst. Als bei den Säugetieren noch alle vier Extremitäten ausschließlich zur Fortbewegung dienten, genügten einfache Reflexmuster auf Rückenmarksebene dazu, den harmonischen Laufrhythmus zu steuern. Bei den Primaten geschieht ein Wandel der Fortbewegung, vom Vierfüßler zum Klettertier. Damit kommt es zu einer Umkonstruktion der vorderen Extremitäten, die zu Greifinstrumenten werden. Das alte Bewegungsmuster der Vierfüßler ist damit überfordert, aber die Großhirnrinde kann sich durch massives Wachstum den neuen Anforderungen der Handmotorik anpassen. ⓘ
Zusätzlich ist bei den Säugern das Kleinhirn in Verbindung mit dem Gleichgewichtsorgan für die Ausführung komplizierter Bewegungsabläufe in das motorische System integriert. Aufrechtes Laufen auf zwei Beinen ist ohne diese Hirnstruktur nicht möglich. Die Zusammenarbeit zwischen Cortex und Kleinhirn lässt sich am Beispiel des Radfahrens so erklären: Die Entscheidung über Rechtskurve oder Bremsvorgang trifft der Cortex, während die Feinarbeit der Gewichtsverlagerung und viele automatische Bewegungsimpulse im Kleinhirn bearbeitet werden. ⓘ
Bei den Affen hat sich die Stellung der Augen im Gesichtsfeld so geändert, dass immer ein räumliches Bild der Umwelt gesehen wird. Für die zentrale Auswertung der binokularen Bilder müssen neue Analysatoren in das System integriert werden, und auch dabei erweist sich die Großhirnrinde als anpassungsfähiges Integrationszentrum mit riesigem Speichervermögen für komplexe Information. ⓘ
Mit dieser Ausstattung war spätestens Homo erectus für den aufrechten Gang in der Savanne gut gerüstet und konnte den Geruchssinn zu Gunsten der Fernsinne (Augen und Ohren) vernachlässigen. Der Cortex passte sich seinen neuen Anforderungen an, indem er seine Fläche durch Faltenbildung vergrößerte. ⓘ
So weit ist das biologische Standardwissen detailliert erforscht und beweist, dass die Großhirnrinde von Anfang an für die Herstellung einer ganzheitlich vereinigten Projektion aller Umweltsignale und einer daraus basierenden Verhaltenssteuerung spezialisiert war und diese Aufgabe in der Evolution immer stärker ausdehnen konnte. Ein bisher noch unverstandener Speichermechanismus ist verantwortlich für die Gedächtnisfunktion dieser Integrationszentrale, die den Lebewesen neben der starren, genetischen Anpassung eine flexible Anpassung an beliebige neue Situationen ermöglicht. ⓘ
Die ersten Menschen hatten mit diesem Gedächtnisorgan und einem verbesserten Kehlkopf die Grundlage für die Verfeinerung der äffischen Laut- und Gebärdensprache. Die veränderte Daumenstellung erleichterte den Gebrauch von Werkzeugen und sorgte für weitere Ausdehnung der Hirnrindentätigkeit. ⓘ
Schon bei der Herstellung von Faustkeilen mit scharfen Klingen ergab sich eine Aufgabenteilung für die beiden Hände, indem eine Hand zum Festhalten und die zweite Hand für gestaltende Feinarbeiten bevorzugt wurden. Viele Tätigkeiten mit Werkzeugen fördern eine differenzierte Spezialisierung der Hände, und spätestens beim systematischen Training des Schreibens ist eine dominante Hand kaum noch zu vermeiden. ⓘ
Dementsprechend unterscheiden sich die beiden Seiten der Hirnrinde im Lauf der Evolution und der individuellen Entwicklung zunehmend, und nur auf der Seite der schreibenden Hand wird zusammen mit den Buchstabenverbindungen auch die Artikulation der Sprache gründlich trainiert. Weil die Nervenbahn des rechten, schreibenden Armes im linken Cortex beginnt, liegen auch die Sprachzentren im linken Großhirn, das deshalb als die dominante Hemisphäre bezeichnet wird. ⓘ
Die Evolution des Cortex ist nachvollziehbar. Es fehlt nur noch eine wissenschaftlich einleuchtende Erklärung für die Leistungsfähigkeit, die sich in den grauen Falten unter der Schädeldecke als Gedächtnis und Bewusstsein erleben und in Sprache ausdrücken lässt. ⓘ