Moas
| Moa | |
|---|---|
.jpg)
| |
| Skelett des Nordinsel-Riesenmoas | |
| Wissenschaftliche Klassifizierung | |
| Königreich: | Tierreich (Animalia) |
| Stamm: | Chordata |
| Klasse: | Aves |
| Infraklasse: | Palaeognathae |
| Klade: | Notopalaeognathae |
| Ordnung: | †Dinornithiformes Bonaparte, 1853 |
| Typusart | |
| †Dinornis novaezealandiae Owen, 1843
| |
| Untergruppen | |
|
Siehe Text | |
| Artenvielfalt | |
| 6 Gattungen, 9 Arten | |
| Synonyme | |
| |
Moa (Ordnung Dinornithiformes) waren neun Arten (in sechs Gattungen) von heute ausgestorbenen flugunfähigen Vögeln, die in Neuseeland endemisch waren. Die beiden größten Arten, Dinornis robustus und Dinornis novaezelandiae, erreichten mit ausgestrecktem Hals eine Höhe von etwa 3,6 m und wogen etwa 230 kg, während die kleinste Art, der Buschmoa (Anomalopteryx didiformis), etwa so groß wie ein Truthahn war. Die Schätzungen der Moa-Population zur Zeit der Besiedlung Neuseelands durch die Polynesier um 1300 schwanken zwischen 58.000 und etwa 2,5 Millionen. ⓘ
Moa werden traditionell zur Gruppe der Laufvögel gezählt. Genetische Untersuchungen haben jedoch ergeben, dass ihre engsten Verwandten die flugfähigen südamerikanischen Tinamus sind, die früher als Schwestergruppe der Laufvögel angesehen wurden. Die neun Moa-Arten waren die einzigen flügellosen Vögel, denen sogar die rudimentären Flügel fehlten, die alle anderen Laufvögel haben. Bis zur Ankunft der Māori waren sie die größten Landtiere und die wichtigsten Pflanzenfresser in Neuseelands Wald-, Buschland- und subalpinen Ökosystemen und wurden nur vom Haast-Adler gejagt. Die Moa starben innerhalb von 100 Jahren nach der Besiedlung Neuseelands durch den Menschen aus, was hauptsächlich auf die Überjagung durch die Māori zurückzuführen ist. ⓘ
| Moas ⓘ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Zwei Moas werden von einem Haastadler attackiert. | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Dinornithiformes | ||||||||||
| Bonaparte, 1853 |
.png)
Etymologie
Das Wort Moa ist eine polynesische Bezeichnung für Hausgeflügel. Der Name war bei den Māori zur Zeit des Kontakts mit den Europäern noch nicht gebräuchlich, wahrscheinlich weil der Vogel, den er beschrieb, schon seit einiger Zeit ausgestorben war und es kaum noch traditionelle Geschichten über ihn gab. Die früheste Erwähnung des Namens stammt von den Missionaren William Williams und William Colenso im Januar 1838; Colenso spekulierte, dass die Vögel riesigen Hühnern geähnelt haben könnten. Im Jahr 1912 behauptete der Māori-Häuptling Urupeni Pūhara, der traditionelle Name des Moas sei "te kura" (der rote Vogel). ⓘ
Beschreibung

Moa-Skelette wurden traditionell in aufrechter Position rekonstruiert, um eine beeindruckende Körpergröße zu erreichen, aber die Analyse ihrer Wirbelgelenke zeigt, dass sie ihren Kopf wahrscheinlich nach vorne trugen, ähnlich wie eine Kiwi. Die Wirbelsäule war eher an der Rückseite des Kopfes als an der Basis befestigt, was auf eine horizontale Ausrichtung hindeutet. Dies hätte es ihnen ermöglicht, auf niedriger Vegetation zu grasen, während sie gleichzeitig in der Lage waren, ihren Kopf zu heben und Bäume zu durchstöbern, wenn es nötig war. Dies hat dazu geführt, dass die Größe größerer Moa neu überdacht wurde. Die Felszeichnungen der Māori zeigen jedoch Moas oder moaähnliche Vögel (wahrscheinlich Gänse oder Adzebills) mit aufrechtem Hals, was darauf hindeutet, dass Moas durchaus in der Lage waren, beide Halshaltungen einzunehmen. ⓘ
Es gibt keine Aufzeichnungen darüber, welche Laute Moas von sich gaben, obwohl fossile Funde eine gewisse Vorstellung von ihren Rufen vermitteln können. Die Luftröhre der Moa wurde von vielen kleinen Knochenringen gestützt, die als Tracheenringe bekannt sind. Die Ausgrabung dieser Ringe aus artikulierten Skeletten hat gezeigt, dass mindestens zwei Moa-Gattungen (Euryapteryx und Emeus) verlängerte Luftröhren hatten, d. h. ihre Luftröhren waren bis zu 1 m lang und bildeten eine große Schleife in der Körperhöhle. Sie sind die einzigen Laufvögel, von denen dieses Merkmal bekannt ist, das auch bei mehreren anderen Vogelgruppen vorkommt, darunter Schwäne, Kraniche und Perlhühner. Dieses Merkmal ist mit tiefen, resonanten Lauten verbunden, die weite Strecken zurücklegen können. ⓘ
Evolutionäre Beziehungen

Die engsten Verwandten des Moas sind kleine südamerikanische Landvögel, die Tinamous, die fliegen können. Früher hielt man den Kiwi, den australischen Emu und den Kasuar für die engsten Verwandten des Moas. ⓘ
Obwohl im späten 19. und frühen 20. Jahrhundert Dutzende von Arten beschrieben wurden, basierten viele auf Teilskeletten und erwiesen sich als Synonyme. Derzeit sind 11 Arten offiziell anerkannt, obwohl neuere Studien, bei denen alte DNA aus Knochen in Museumssammlungen verwendet wurde, darauf hindeuten, dass es innerhalb einiger dieser Arten unterschiedliche Abstammungslinien gibt. Ein Faktor, der in der Moa-Taxonomie für viel Verwirrung gesorgt hat, ist die intraspezifische Variation der Knochengrößen zwischen den Eiszeiten und den Zwischeneiszeiten (siehe Bergmannsche Regel und Allensche Regel) sowie der Geschlechtsdimorphismus, der bei mehreren Arten zu beobachten ist. Dinornis scheint den ausgeprägtesten Sexualdimorphismus aufzuweisen, wobei die Weibchen bis zu 150 % so groß und 280 % so schwer sind wie die Männchen - so viel größer, dass sie bis 2003 als eigene Arten eingestuft wurden. Eine Studie von 2009 zeigte, dass Euryapteryx curtus und E. gravis Synonyme sind. Eine Studie aus dem Jahr 2010 erklärte die Größenunterschiede zwischen ihnen als Sexualdimorphismus. In einer morphologischen Studie von 2012 wurden sie stattdessen als Unterarten interpretiert. ⓘ
Analysen alter DNA haben ergeben, dass es in mehreren Moa-Gattungen eine Reihe von kryptischen Evolutionslinien gibt. Diese können möglicherweise als Arten oder Unterarten klassifiziert werden; Megalapteryx benhami (Archey) wird mit M. didinus (Owen) synonymisiert, da die Knochen beider Arten alle wesentlichen Merkmale gemeinsam haben. Die Größenunterschiede lassen sich durch eine Nord-Süd-Grenze in Verbindung mit zeitlichen Schwankungen erklären, so dass die Exemplare während der Otiran-Glazialzeit (der letzten Eiszeit in Neuseeland) größer waren. Eine ähnliche zeitliche Größenvariation ist für Pachyornis mappini von der Nordinsel bekannt. Einige der anderen Größenunterschiede bei Moa-Arten lassen sich wahrscheinlich durch ähnliche geografische und zeitliche Faktoren erklären. ⓘ
Die frühesten Moa-Überreste stammen aus der miozänen Saint Bathans Fauna. Sie sind durch mehrere Eierschalen und Elemente der Hinterbeine bekannt und repräsentieren mindestens zwei bereits recht große Arten. ⓘ
Klassifizierung
Taxonomie

_601344.jpg)
Die derzeit anerkannten Gattungen und Arten sind:
- Ordnung †Dinornithiformes (Gadow 1893) Ridgway 1901 [Dinornithes Gadow 1893; Immanes Newton 1884] (Moa)
- Familie Dinornithidae Owen 1843 [Palapteryginae Bonaparte 1854; Palapterygidae Haast 1874; Dinornithnideae Stejneger 1884] (Riesenmoa)
- Gattung Dinornis
- Nordinsel-Riesenmoa, Dinornis novaezealandiae (Nordinsel, Neuseeland)
- Südinsel-Riesenmoa, Dinornis robustus (Südinsel, Neuseeland)
- Gattung Dinornis
- Familie Emeidae (Bonaparte 1854) [Emeinae Bonaparte 1854; Anomalopterygidae Oliver 1930; Anomalapteryginae Archey 1941] (Kleiner Moa)
- Gattung Anomalopteryx
- Buschmoa, Anomalopteryx didiformis (Nord- und Südinsel, Neuseeland)
- Gattung Emeus
- Östlicher Moa, Emeus crassus (Südinsel, Neuseeland)
- Gattung Euryapteryx
- Breitschnabelmoa, Euryapteryx curtus (Nord- und Südinsel, Neuseeland)
- Gattung Pachyornis
- Schwerfüßiger Moa, Pachyornis elephantopus (Südinsel, Neuseeland)
- Mantelsumpfhuhn, Pachyornis geranoides (Nordinsel, Neuseeland)
- Haubenmoa, Pachyornis australis (Südinsel, Neuseeland)
- Gattung Anomalopteryx
- Familie Megalapterygidae
- Gattung Megalapteryx
- Hochlandmoa, Megalapteryx didinus (Südinsel, Neuseeland) ⓘ
- Gattung Megalapteryx
- Familie Dinornithidae Owen 1843 [Palapteryginae Bonaparte 1854; Palapterygidae Haast 1874; Dinornithnideae Stejneger 1884] (Riesenmoa)
Zwei unbenannte Arten aus der Fauna von Saint Bathans. ⓘ
Phylogenie
Da die Moa zu den flugunfähigen Vögeln gehören, die keine Flügelknochen besitzen, wurde die Frage aufgeworfen, wie und woher sie nach Neuseeland kamen. Es gibt viele Theorien über die Ankunft und Ausbreitung der Moa in Neuseeland, aber die neueste Theorie besagt, dass sie vor etwa 60 Millionen Jahren (Mya) nach Neuseeland kamen und sich von der "basalen" (siehe unten) Moa-Art Megalapteryx vor etwa 5,8 Mya abspalteten, anstatt der von Baker et al. (2005) vorgeschlagenen Spaltung vor 18,5 Mya. Dies bedeutet nicht zwangsläufig, dass es zwischen der Ankunft 60 Mya und der basalen Spaltung 5,8 Mya keine Artenbildung gab, aber es fehlen fossile Aufzeichnungen, und höchstwahrscheinlich existierten die frühen Moa-Linien, starben aber vor der basalen Spaltung 5,8 Mya aus. Das Vorhandensein von Arten aus dem Miozän lässt mit Sicherheit darauf schließen, dass die Diversifizierung der Moa vor der Spaltung zwischen Megalapteryx und den anderen Taxa begann. ⓘ
Das oligozäne Ertränkungsmaximum, das sich um 22 Mya ereignete, als nur 18 % des heutigen Neuseelands über dem Meeresspiegel lagen, ist für die Moa-Radiation sehr wichtig. Da die Spaltung der Moa-Basis erst vor so kurzer Zeit (5,8 Mya) stattfand, wurde argumentiert, dass die Vorfahren der Moa-Linien des Quartärs während des oligozänen Ertränkungsmaximums nicht sowohl auf der Süd- als auch auf der Nordinsel zu finden gewesen sein können. Dies bedeutet nicht, dass es vorher keine Moa auf der Nordinsel gab, sondern dass nur die Moa von der Südinsel überlebten, weil nur die Südinsel über dem Meeresspiegel lag. Bunce et al. (2009) argumentierten, dass die Vorfahren der Moa auf der Südinsel überlebten und dann etwa 2 My später die Nordinsel wieder besiedelten, als die beiden Inseln nach 30 My Trennung wieder zusammenfanden. Das Vorkommen miozäner Moa in der Fauna von Saint Bathans scheint darauf hinzudeuten, dass diese Vögel bald nach dem oligozänen Ertrinkungsereignis an Größe zunahmen, wenn sie überhaupt davon betroffen waren. ⓘ
Bunce et al. kamen außerdem zu dem Schluss, dass die hochkomplexe Struktur der Moa-Stammbäume durch die Bildung der Südalpen um 6 Mya und die Fragmentierung des Lebensraums auf beiden Inseln infolge der pleistozänen Gletscherzyklen, des Vulkanismus und der Landschaftsveränderungen verursacht wurde. Das nachstehende Kladogramm ist eine von Mitchell (2014) erstellte Phylogenie der Palaeognathae mit einigen Kladennamen nach Yuri et al. (2013). Es zeigt die Position der Moa (Dinornithiformes) innerhalb des größeren Kontextes der Vögel mit "altem Maul" (Palaeognathae):
ⓘ| Palaeognathae |
| ||||||||||||||||||||||||||||||||||||||||||
Das nachstehende Kladogramm zeigt eine detailliertere Phylogenie auf Artniveau des Moa-Zweiges (Dinornithiformes) der oben gezeigten "altkieferartigen" Vögel (Palaeognathae):
ⓘ| †Dinornithiformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verbreitung und Lebensraum
Analysen fossiler Moa-Knochen haben detaillierte Daten über die Lebensraumpräferenzen einzelner Moa-Arten geliefert und charakteristische regionale Moa-Faunen offenbart: ⓘ
Südinsel

Auf der Südinsel wurden zwei Hauptfaunen festgestellt:
- Die Fauna der niederschlagsreichen Buchenwälder (Nothofagus) an der Westküste, zu der Anomalopteryx didiformis (Buschmoa) und Dinornis robustus (Südinsel-Riesenmoa) gehören, und
- Die Fauna der trockenen Regenwälder und Buschländer östlich der Südalpen, zu der Pachyornis elephantopus (Schwerfußmoa), Euryapteryx gravis, Emeus crassus und Dinornis robustus gehörten.
Eine "subalpine Fauna" könnte den weit verbreiteten D. robustus und die beiden anderen Moa-Arten umfassen, die auf der Südinsel vorkamen:
- Pachyornis australis, die seltenste Moaart, die einzige Moaart, die noch nicht in Māori-Middens gefunden wurde. Seine Knochen wurden in Höhlen in den nordwestlichen Bezirken Nelson und Karamea (z. B. Honeycomb Hill Cave) sowie an einigen Stellen im Bezirk Wānaka gefunden.
- Megalapteryx didinus, weiter verbreitet, auch "Hochlandmoa" genannt, weil seine Knochen häufig in der subalpinen Zone gefunden werden. Er kam jedoch auch bis zum Meeresspiegel vor, wo es geeignetes steiles und felsiges Gelände gab (z. B. Punakaiki an der Westküste und Zentral-Otago). Ihr Verbreitungsgebiet in den Küstengebieten ist eher unklar, aber sie kam zumindest an mehreren Orten vor, wie z. B. auf Kaikōura, der Otago-Halbinsel und Karitane. ⓘ

Nordinsel
Über die Paläofauna der Nordinsel ist deutlich weniger bekannt, da es im Vergleich zur Südinsel nur wenige Fossilienfunde gibt, aber das Grundmuster der Beziehungen zwischen Moa und Lebensraum war dasselbe. Die Südinsel und die Nordinsel hatten einige Moa-Arten gemeinsam (Euryapteryx gravis, Anomalopteryx didiformis), aber die meisten Arten kamen nur auf einer Insel vor, was die Divergenz über mehrere tausend Jahre hinweg widerspiegelt, nachdem der niedrigere Meeresspiegel in der Eiszeit eine Landbrücke über die Cookstraße geschaffen hatte. ⓘ
Auf der Nordinsel dominierten Dinornis novaezealandiae und Anomalopteryx didiformis in regenreichen Waldlebensräumen, ein ähnliches Muster wie auf der Südinsel. Die anderen auf der Nordinsel vorkommenden Moa-Arten (Euryapteryx gravis, E. curtus und Pachyornis geranoides) bewohnten eher trockenere Wald- und Strauchland-Lebensräume. P. geranoides kam auf der gesamten Nordinsel vor. Die Verbreitungsgebiete von E. gravis und E. curtus schlossen sich fast gegenseitig aus, wobei E. gravis nur an Küstenstandorten in der südlichen Hälfte der Nordinsel gefunden wurde. ⓘ
Lebensweise und Ökologie

Etwa acht Moa-Fährten mit versteinerten Moa-Fußabdrücken in Flussschlamm wurden auf der Nordinsel gefunden, darunter Waikanae Creek (1872), Napier (1887), Manawatū River (1895), Marton (1896), Palmerston North (1911) (siehe Foto links), Rangitīkei River (1939) und unter Wasser im Lake Taupō (1973). Die Analyse der Abstände dieser Spuren deutet auf Laufgeschwindigkeiten zwischen 3 und 5 km/h (1,75-3 mph) hin. ⓘ
Ernährung

Ihre Ernährung wurde aus dem fossilen Inhalt ihrer Mägen und Koprolithen sowie indirekt durch die morphologische Analyse von Schädel und Schnabel und die Analyse stabiler Isotope ihrer Knochen abgeleitet. Moa ernährten sich von einer Reihe von Pflanzenarten und Pflanzenteilen, darunter faserige Zweige und Blätter von niedrigen Bäumen und Sträuchern. Der Schnabel von Pachyornis elephantopus glich einer Gartenschere und konnte die faserigen Blätter des neuseeländischen Flachses (Phormium tenax) und Zweige mit einem Durchmesser von mindestens 8 mm abschneiden. ⓘ
Der Moa füllte die ökologische Nische, die in anderen Ländern von großen, grasenden Säugetieren wie Antilopen und Lamas besetzt wurde. Einige Biologen sind der Meinung, dass sich eine Reihe von Pflanzenarten entwickelt haben, um dem Moa-Verbiss zu entgehen. Spaltpflanzen wie Pennantia corymbosa (kaikōmako) mit kleinen Blättern und einem dichten Astgeflecht und Pseudopanax crassifolius (horoeka oder Lanzenholz) mit zähen jungen Blättern sind mögliche Beispiele für Pflanzen, die sich auf diese Weise entwickelt haben. ⓘ
Wie viele andere Vögel verschluckten auch die Moa Magensteine (Gastrolithen), die in ihrem Muskelmagen verbleiben und eine Mahlwirkung haben, die es ihnen ermöglicht, grobes Pflanzenmaterial zu fressen. Bei diesen Steinen handelte es sich in der Regel um glatte, abgerundete Quarzkiesel, aber auch Steine mit einer Länge von mehr als 110 Millimetern wurden in erhaltenen Mägen von Moas gefunden. Dinornis-Mägen konnten oft mehrere Kilogramm Steine enthalten. Wahrscheinlich haben die Mäuse bei der Auswahl der Kaumagensteine eine gewisse Selektivität an den Tag gelegt und die härtesten Kiesel ausgewählt. ⓘ
Fortpflanzung
Die als Euryapteryx curtus / E. exilis, Emeus huttonii / E. crassus und Pachyornis septentrionalis / P. mappini bezeichneten Moa-Artenpaare werden seit langem als Männchen bzw. Weibchen angesehen. Dies wurde durch die Analyse geschlechtsspezifischer genetischer Marker von aus Knochenmaterial gewonnener DNA bestätigt. ⓘ
Vor 2003 wurden beispielsweise drei Arten von Dinornis anerkannt: Südinsel-Riesenmoa (D. robustus), Nordinsel-Riesenmoa (D. novaezealandiae) und Schlankmoa (D. struthioides). Die DNA-Analyse ergab jedoch, dass alle D. struthioides Männchen und alle D. robustus Weibchen waren. Daher wurden die drei Dinornis-Arten in zwei Arten umgewandelt, von denen jeweils eine früher auf der Nordinsel (D. novaezealandiae) und der Südinsel (D. robustus) Neuseelands vorkam; D. robustus umfasst jedoch drei verschiedene genetische Linien und könnte, wie oben erläutert, als mehrere Arten klassifiziert werden. ⓘ
Die Untersuchung von Wachstumsringen in den Rindenknochen der Moa hat ergeben, dass diese Vögel, wie viele andere große endemische Vögel Neuseelands, K-selektiert wurden. Sie zeichnen sich durch eine geringe Fruchtbarkeit und eine lange Reifezeit aus, die bis zum Erreichen der Erwachsenengröße etwa 10 Jahre dauert. Die großen Dinornis-Arten brauchten genauso lange, um die Erwachsenengröße zu erreichen, wie die kleinen Moa-Arten, und hatten daher ein schnelles Skelettwachstum während ihrer Jugendjahre. ⓘ
Es wurden keine Beweise dafür gefunden, dass Moas koloniale Nester angelegt haben. Das Nisten von Moas wird häufig aus Anhäufungen von Eierschalenfragmenten in Höhlen und Felsunterkünften abgeleitet, für die Nester selbst gibt es nur wenige Belege. Bei Ausgrabungen von Felsunterkünften auf der östlichen Nordinsel in den 1940er Jahren wurden Moa-Nester gefunden, die als "kleine, offensichtlich in den weichen, trockenen Bimsstein geritzte Vertiefungen" beschrieben wurden. Auch in der Region Central Otago auf der Südinsel wurde Nistmaterial von Moas in Felsen gefunden, wo das trockene Klima Pflanzenmaterial zum Bau der Nistplattform konserviert hat (einschließlich Zweige, die von Moa-Schnäbeln abgeschnitten wurden). Samen und Pollen in Moa-Koprolithen, die unter dem Nistmaterial gefunden wurden, belegen, dass die Nistzeit im späten Frühjahr bis zum Sommer war. ⓘ
Fragmente von Moa-Eiern werden häufig in archäologischen Stätten und Sanddünen an der neuseeländischen Küste gefunden. Sechsunddreißig ganze Moa-Eier sind in Museumssammlungen vorhanden und variieren stark in ihrer Größe (von 120-240 Millimeter Länge und 91-178 Millimeter Breite). Die Außenfläche der Moa-Eierschale ist durch kleine, schlitzförmige Poren gekennzeichnet. Die Eier der meisten Moa-Arten sind weiß, die des Hochland-Moas (Megalapteryx didinus) sind jedoch blaugrün. ⓘ
In einer Studie von Huynen et al. aus dem Jahr 2010 wurde festgestellt, dass die Eier bestimmter Arten zerbrechlich sind und die Schale nur etwa einen Millimeter dick ist: "Unerwartet zeigte sich, dass mehrere dünnschalige Eier auch zu den schwersten Moas der Gattungen Dinornis, Euryapteryx und Emeus gehörten, was sie unseres Wissens nach zu den zerbrechlichsten aller bisher gemessenen Vogeleier macht. Die geschlechtsspezifische DNA, die an den Außenflächen der Eierschalen von Dinornis- und Euryapteryx-Arten gefunden wurde, deutet außerdem darauf hin, dass diese sehr dünnen Eier wahrscheinlich von den leichteren Männchen bebrütet wurden. Die dünne Beschaffenheit der Eierschalen dieser größeren Moa-Arten, selbst wenn sie vom Männchen bebrütet wurden, deutet darauf hin, dass die Eier dieser Arten häufig zerbrochen wurden, wenn die typische Kontaktmethode für die Bebrütung von Vogeleiern angewandt wurde." Trotz des Aussterbens dieses Vogels konnte das Genom des Moas dank der großen Menge an DNA aus den geborgenen fossilen Eiern sequenziert werden. ⓘ

Das Skelett eines weiblichen Hochlandmoas mit einem Ei, das sich in der Beckenhöhle befindet, im Otago Museum
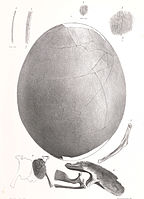
Ein Ei und Fragmente eines Embryos von Emeus crassus

Wiederherstellung eines Hochlandmoas ⓘ



Moas waren ausschließlich Pflanzenfresser. Durch Untersuchungen der Muskelmägen bei besonders gut erhaltenen Moa-Fossilien konnte man feststellen, dass Dinornis offenbar hauptsächlich Zweige abweideten, während Emeus und Euryapteryx weichere Kost wie Blätter und Früchte zu sich nahmen. Von Vertretern anderer Gattungen sind keine Mageninhalte bekannt, jedoch bis zu fünf Zentimeter große Magensteine (Gastrolithen). Für einen Anteil tierischer Nahrung gibt es keine Anhaltspunkte. ⓘ
Auch über die Laute, die Moas von sich gaben, konnte man durch die Untersuchung eines mumifizierten Euryapteryx Klarheit erlangen. Bei diesem bildet die Luftröhre eine 1,20 Meter lange Schleife, eine Struktur, die man ähnlich beim Trompeterschwan findet. Ein solches Organ ermöglichte dem Vogel, sehr laute und weit tragende Rufe zu erzeugen. Ob andere Moa-Gattungen vergleichbare Vorrichtungen hatten, ist im Moment noch Spekulation. ⓘ
Vor der Ankunft des Menschen war der Haastadler der einzige Feind der Moas. Für ihn waren vor allem die kleinen und mittelgroßen Arten die Hauptbeute. Doch auch die riesigen Weibchen der beiden Dinornis-Arten fielen dem gigantischen Greifvogel gelegentlich zum Opfer. Dies weiß man von der Untersuchung der Überreste verschiedener Moas, die schwere Verletzungsspuren des Beckens aufweisen. Sie deuten darauf hin, dass der Adler seine Beute von hinten attackierte. Die Beckenknochen wurden von den Adlerkrallen regelrecht durchstochen. ⓘ
Vormenschliche Wälder
Untersuchungen der angesammelten getrockneten Vegetation in der vormenschlichen Periode des mittleren bis späten Holozäns deuten auf ein niedriges Sophora microphylla- oder Kōwai-Wald-Ökosystem in Zentral-Otago hin, das von Moas sowohl als Nistmaterial als auch als Nahrung genutzt und möglicherweise erhalten wurde. Als die europäischen Siedler in den 1850er Jahren in dieses Gebiet kamen, gab es weder Wälder noch Moa. ⓘ
Beziehung zum Menschen
Aussterben
Vor der Ankunft der menschlichen Siedler war der einzige Raubtier der Moa der riesige Haast-Adler. Neuseeland war 80 Millionen Jahre lang isoliert gewesen und hatte vor der Ankunft des Menschen nur wenige Raubtiere, was nicht nur bedeutete, dass das Ökosystem extrem anfällig für Störungen durch fremde Arten war, sondern auch, dass die einheimischen Arten schlecht gerüstet waren, um mit menschlichen Raubtieren fertig zu werden. ⓘ
Die Ankunft der Polynesier erfolgte irgendwann vor 1300, und alle Moa-Gattungen wurden bald durch die Jagd und in geringerem Maße durch die Verringerung des Lebensraums infolge der Abholzung der Wälder zum Aussterben gebracht. Um 1445 waren alle Moas ausgestorben, ebenso wie der Haast-Adler, der auf sie als Nahrung angewiesen war. Jüngste Forschungen, bei denen Kohlenstoff-14-Datierungen von Mülldeponien verwendet wurden, deuten darauf hin, dass die Ereignisse, die zum Aussterben führten, weniger als hundert Jahre dauerten und nicht, wie früher angenommen, eine mehrere hundert Jahre dauernde Ausbeutungsperiode. ⓘ
Einige Autoren haben spekuliert, dass einige wenige Megalapteryx didinus in abgelegenen Ecken Neuseelands bis ins 18. oder sogar 19. Jahrhundert überlebt haben könnten, aber diese Ansicht wird nicht allgemein akzeptiert. Einige Māori-Jäger behaupteten noch in den 1770er Jahren, den Moa zu jagen; diese Berichte bezogen sich jedoch möglicherweise nicht auf die Jagd nach den Vögeln selbst, sondern auf ein inzwischen verloren gegangenes Ritual der Südinsulaner. Walfänger und Robbenfänger erinnerten sich daran, monströse Vögel an der Küste der Südinsel gesehen zu haben, und in den 1820er Jahren behauptete ein Mann namens George Pauley unbestätigt, einen Moa in der Region Otago in Neuseeland gesehen zu haben. ⓘ
Eine Expedition unter Leutnant A. Impey in den 1850er Jahren berichtete von zwei emu-ähnlichen Vögeln an einem Berghang auf der Südinsel; ein Bericht des Nelson Examiner aus dem Jahr 1861 berichtete von Fußabdrücken von drei Zehen, die 36 cm groß waren und von einem Vermessungstrupp zwischen Tākaka und Riwaka gefunden wurden; und schließlich veröffentlichte der Otago Witness 1878 einen weiteren Bericht eines Bauern und seines Hirten. Eine 80-jährige Frau, Alice McKenzie, behauptete 1959, sie habe 1887 im Fiordland-Busch einen Moa gesehen, und noch einmal an einem Fiordland-Strand, als sie 17 Jahre alt war. Sie behauptete, auch ihr Bruder habe bei einer anderen Gelegenheit einen Moa gesehen. ⓘ
Überlebende Überreste

Joel Polack, ein Händler, der von 1834 bis 1837 an der Ostküste der Nordinsel lebte, berichtete 1838, dass ihm "mehrere große fossile Verknöcherungen" gezeigt wurden, die in der Nähe des Mount Hikurangi gefunden worden waren. Er war sich sicher, dass es sich dabei um die Knochen einer Emu- oder Straußenart handelte, und bemerkte, dass "die Eingeborenen hinzufügen, dass sie in längst vergangenen Zeiten die Überlieferung erhielten, dass es sehr große Vögel gegeben habe, aber der Mangel an tierischer Nahrung sowie die einfache Methode, sie zu fangen, ihre Ausrottung verursacht habe". Polack merkte ferner an, dass er von den Māori Berichte erhalten habe, wonach es in abgelegenen Teilen der Südinsel noch eine "Art Struthio" gebe. ⓘ
Dieffenbach bezieht sich auch auf ein Fossil aus der Gegend von Mt. Hikurangi und vermutet, dass es zu "einem Vogel gehört, der jetzt ausgestorben ist und von den Eingeborenen Moa (oder Movie) genannt wird". Movie" ist der erste transkribierte Name für diesen Vogel. Im Jahr 1839 erhielt John W. Harris, ein Flachshändler aus Poverty Bay, der sich für Naturgeschichte begeisterte, von einem Māori, der es an einem Flussufer gefunden hatte, ein Stück eines ungewöhnlichen Knochens. Er zeigte das 15 cm lange Knochenfragment seinem Onkel John Rule, einem Chirurgen aus Sydney, der es an Richard Owen schickte, der zu dieser Zeit am Hunterian Museum des Royal College of Surgeons in London tätig war. ⓘ

Owen rätselte fast vier Jahre lang über das Fragment. Er stellte fest, dass es zum Oberschenkelknochen eines großen Tieres gehörte, aber es war untypisch hell und wabenförmig. Owen verkündete einer skeptischen wissenschaftlichen Gemeinschaft und der Welt, dass es von einem riesigen, ausgestorbenen Vogel wie dem Strauß stammte, und nannte es Dinornis. Seine Schlussfolgerung wurde in einigen Kreisen belächelt, erwies sich jedoch als richtig, als in der Folgezeit im ganzen Land beträchtliche Mengen von Moa-Knochen entdeckt wurden, die ausreichten, um die Skelette der Vögel zu rekonstruieren. ⓘ
Im Juli 2004 stellte das Natural History Museum in London das Moa-Knochenfragment aus, das Owen als Erster untersucht hatte, um sein 200-jähriges Bestehen zu feiern und um an Owen als Gründer des Museums zu erinnern. ⓘ

Seit der Entdeckung der ersten Moa-Knochen in den späten 1830er Jahren wurden Tausende weiterer Knochen gefunden. Sie kommen in einer Reihe von Sedimentablagerungen des späten Quartärs und Holozäns vor, sind aber am häufigsten in drei Haupttypen von Fundorten anzutreffen: Höhlen, Dünen und Sümpfen. ⓘ

Knochen werden häufig in Höhlen oder Tomo (das Māori-Wort für Doline oder Sinkloch, das oft für Fallgruben oder vertikale Höhlenschächte verwendet wird) gefunden. Die beiden Hauptursachen für die Ablagerung der Moa-Knochen an solchen Orten sind Vögel, die in die Höhle eingedrungen sind, um zu nisten oder vor schlechtem Wetter zu fliehen, und dann in der Höhle gestorben sind, oder Vögel, die in einen vertikalen Schacht gefallen sind und nicht mehr entkommen konnten. Moa-Knochen (und die Knochen anderer ausgestorbener Vögel) wurden in Höhlen in ganz Neuseeland gefunden, insbesondere in den Kalkstein-/Marmorgebieten im Nordwesten von Nelson, Karamea, Waitomo und Te Anau. ⓘ
Moa-Knochen und Eierschalenfragmente kommen manchmal in aktiven Küstendünen vor, wo sie von Paläosolen erodieren und sich in "Ausbrüchen" zwischen Dünenkämmen konzentrieren können. Viele dieser Moa-Knochen stammen aus der Zeit vor der Besiedlung durch den Menschen, obwohl einige von Māori-Midden stammen, die häufig in Dünen in der Nähe von Häfen und Flussmündungen vorkommen (z. B. die großen Moa-Jäger-Fundstellen in Shag River, Otago, und Wairau Bar, Marlborough). ⓘ
In den Sümpfen Neuseelands wurden dicht verstreute Moa-Knochen gefunden. Das bekannteste Beispiel ist das Pyramid Valley in Nord-Canterbury, wo Knochen von mindestens 183 einzelnen Moas ausgegraben wurden, hauptsächlich von Roger Duff vom Canterbury Museum. Es gibt viele Erklärungen für die Entstehung dieser Ablagerungen, die von giftigem Quellwasser bis hin zu Überschwemmungen und Waldbränden reichen. Die gegenwärtig akzeptierte Erklärung ist jedoch, dass sich die Knochen langsam über Tausende von Jahren angesammelt haben, und zwar von Vögeln, die in die Sümpfe eingedrungen sind, um zu fressen, und sich in den weichen Sedimenten verfangen haben. ⓘ
Viele neuseeländische und internationale Museen besitzen Sammlungen von Moa-Knochen. Das Auckland War Memorial Museum - Tāmaki Paenga Hira verfügt über eine bedeutende Sammlung, und 2018 wurden mehrere Moa-Skelette abgebildet und 3D-gescannt, um die Sammlungen besser zugänglich zu machen. ⓘ
Federn und Weichteile

Es wurden mehrere bemerkenswerte Beispiele von Moa-Überresten gefunden, die Weichgewebe (Muskeln, Haut, Federn) aufweisen, die durch Austrocknung konserviert wurden, wenn der Vogel an einem natürlich trockenen Ort starb (z. B. in einer Höhle, durch die ständig eine trockene Brise weht). Die meisten dieser Exemplare wurden in der semiariden Region Central Otago gefunden, dem trockensten Teil Neuseelands. Dazu gehören:
- Getrocknete Muskeln an den Knochen eines weiblichen Dinornis robustus, die 1864 von Goldgräbern am Tiger Hill im Manuherikia River Valley gefunden wurden (derzeit im Besitz des Yorkshire Museum)
- Mehrere Knochen von Emeus crassus mit anhaftenden Muskeln und eine Reihe von Halswirbeln mit Muskeln, Haut und Federn, die 1870 in der Earnscleugh-Höhle in der Nähe der Stadt Alexandra gefunden wurden (derzeit im Otago Museum)
- Ein artikulierter Fuß eines männlichen D. giganteus mit erhaltener Haut und Fußballen, der 1874 in einer Felsspalte auf der Knobby Range gefunden wurde (derzeit im Besitz des Otago Museums)
- Das Typusexemplar von Megalapteryx didinus, das 1878 in der Nähe von Queenstown gefunden wurde (heute im Natural History Museum, London; siehe Foto des Fußes auf dieser Seite)
- Der Unterschenkel von Pachyornis elephantopus, mit Haut und Muskeln, gefunden 1884 in der Hector Range; (derzeit im Besitz des Zoologischen Instituts der Universität Cambridge)
- Das vollständige gefiederte Bein eines M. didinus aus der Old Man Range von 1894 (derzeit im Besitz des Otago Museums)
- Der Kopf eines M. didinus, der irgendwann vor 1949 in der Nähe von Cromwell gefunden wurde (derzeit im Besitz des Museum of New Zealand). ⓘ
Zwei Exemplare sind von außerhalb der Region Central Otago bekannt:
- Ein vollständiger Fuß von M. didinus, der in den 1980er Jahren in einer Höhle am Mount Owen bei Nelson gefunden wurde (derzeit im Besitz des Museum of New Zealand)
- Ein Skelett von Anomalopteryx didiformis mit Muskeln, Haut und Federbasen, das 1980 in einer Höhle bei Te Anau gefunden wurde. ⓘ

Zusätzlich zu diesen Exemplaren wurden lose Moa-Federn aus Höhlen und Felsunterkünften auf der südlichen Südinsel gesammelt, und anhand dieser Überreste konnte man sich eine Vorstellung vom Gefieder der Moa machen. Das erhaltene Bein von M. didinus aus der Old Man Range zeigt, dass diese Art bis zum Fuß hinunter befiedert war. Dies ist wahrscheinlich eine Anpassung an das Leben in hochgelegenen, verschneiten Umgebungen und wird auch beim Darwin-Rhea beobachtet, der in einem ähnlichen saisonal verschneiten Lebensraum lebt. ⓘ
Die Federn des Moas sind bis zu 23 cm lang, und es wurde von einer Reihe von Farben berichtet, darunter rötlich-braun, weiß, gelblich und violett. Es wurden auch dunkle Federn mit weißen oder cremefarbenen Spitzen gefunden, was darauf hindeutet, dass einige Moa-Arten ein geflecktes Gefieder hatten. ⓘ
Behauptungen über das Überleben

Mindestens seit dem späten 19. Jahrhundert und in jüngster Zeit (1993 und 2008) wird gelegentlich spekuliert, dass es noch Moas geben könnte, insbesondere in der Wildnis von South Westland und Fiordland. Der Bericht von 1993 interessierte zunächst das Department of Conservation, doch das Tier auf einem verschwommenen Foto wurde als Rothirsch identifiziert. Kryptozoologen suchen weiterhin nach ihnen, aber ihre Behauptungen und Beweise (wie z. B. angebliche Fußabdrücke) werden von Experten kaum beachtet und sind pseudowissenschaftlich. Im Jahr 1880 hatte Alice Mackenzie eine Begegnung mit einem großen Vogel, den sie für einen takahē hielt. Als er jedoch in den 1940er Jahren wiederentdeckt wurde und Mackenzie sah, wie er aussah, wusste sie, dass sie etwas anderes gesehen hatte. ⓘ
Die Wiederentdeckung des takahē im Jahr 1948, nachdem seit 1898 keiner mehr gesehen worden war, zeigte, dass seltene Vögel lange Zeit unentdeckt bleiben können. Allerdings ist der takahē ein viel kleinerer Vogel als der Moa und wurde erst wiederentdeckt, nachdem seine Spuren identifiziert worden waren - allerdings wurden noch nie verlässliche Beweise für Moa-Spuren gefunden, und Experten halten ein Überleben des Moas nach wie vor für äußerst unwahrscheinlich, da er über 500 Jahre lang unbemerkt in einer Region leben müsste, die häufig von Jägern und Wanderern besucht wird. ⓘ
Mögliche Wiederbelebung
Die Kreatur wurde häufig als potenzieller Kandidat für eine Wiederbelebung durch Klonen genannt. Sein Kultstatus und die Tatsache, dass er erst vor wenigen hundert Jahren ausgestorben ist und dass es große Mengen an Moa-Überresten gibt, haben dazu geführt, dass er neben Tieren wie dem Dodo oft als Hauptkandidat für seine Ausrottung genannt wird. Der japanische Genetiker Ankoh Yasuyuki Shirota hat erste Arbeiten zur Gewinnung von DNA durchgeführt. ⓘ
Das Interesse an einer möglichen Wiederbelebung des Moas wurde Mitte 2014 weiter geweckt, als der neuseeländische Abgeordnete Trevor Mallard vorschlug, dass die Wiedereinführung einiger kleinerer Moa-Arten innerhalb von 50 Jahren eine realistische Idee sei. Die Idee wurde von vielen belächelt, fand aber bei einigen Naturhistorikern Unterstützung. ⓘ
In Literatur und Kultur

Heinrich Harder porträtierte in den klassischen deutschen Sammelkarten über ausgestorbene und prähistorische Tiere, "Tiere der Urwelt", in den frühen 1900er Jahren Moas, die von Māori gejagt wurden. ⓘ
Allen Curnows Gedicht "The Skeleton of the Great Moa in the Canterbury Museum, Christchurch" wurde 1943 veröffentlicht. ⓘ
Merkmale
Als Urkiefervögel (Palaeognathae) besaßen Moas einen palaeognathen Gaumen, das heißt, sie zeigen im Gegensatz zu den so genannten Neukiefervögeln einen Pterygoid-Palatinum-Komplex (PPC), der aus den Schädelknochen Flügelbein (Pterygoid), Gaumenbein (Palatinum) und Pflugscharbein (Vomer) besteht. Sie glichen anderen Laufvögeln in einer Anzahl anatomischer Merkmale: Ihre Ober- und Unterkiefer endeten jeweils in einem dreiteiligen Hornschnabel (Rhamphotheca), deren mittlerer Kamm flach und von den jeweils seitlichen Schnabelteilen durch Furchen getrennt war. Der Nasalfortsatz des Zwischenkieferbeins war unpaarig und nicht mit dem Nasale verschmolzen. Im Beckengürtel war das Fenster zwischen Darmbein und Sitzbein (Foramen ilioischiadicum) länglich und nach hinten nicht abgeschlossen. Das breite und flache Brustbein war ohne Anzeichen eines Brustbeinkiels (Carina sterni). ⓘ
Durch folgende abgeleitete Merkmale des Skeletts, in denen sich Moas von anderen Laufvögeln unterschieden, ist die Ordnung der Moas (Dinornithiformes) definiert: Flügel fehlten völlig. Vom Schultergürtel lag nur ein verkümmerter Scapulocoracoid ohne Anzeichen eines Schultergelenksockels vor. Das Becken war breit im Bereich der Hüftgelenkpfanne und dahinter. Das Brustbein zeigte gut ausgeprägte seitliche Fortsätze. Das fußwärtige Ende des Schienbeins war mit einem Sehnenkanal ausgestattet, der von einer knöchernen Verbindung überbrückt wurde. Das Laufbein besaß zwei Hypotarsalkämme anstatt eines. Moas besaßen 21 bis 23 Halswirbel, 6 Brustwirbel, 18 Beckenwirbel und 11 Schwanzwirbel, wobei die letzten Schwanzwirbel nicht zu einem Pygostyl verschmolzen waren. Eine Anzahl Schädelmerkmale kommt zu den genannten Charakteristika hinzu. ⓘ
Die meisten Moa-Arten waren kurzbeinig und so groß wie ein Truthahn. Hingegen waren die Weibchen der beiden Arten der Gattung Dinornis die größten Vögel Neuseelands. Ihr Gewicht betrug etwa 180 Kilogramm, nach anderen Schätzungen bis zu 270 Kilogramm. Sie hielten den Kopf nach vorne gestreckt und in Höhe des Rückens oder darunter; dies war der Form der Wirbelsäule geschuldet, die vor den Brustwirbeln abwärts gebogen war, mit dem tiefsten Punkt zwischen den Wirbeln 12 und 16 (vom Schädel aus gezählt); die vorderen Halswirbel waren wiederum so kurz, dass sie kaum zur Erhöhung des Kopfes beitrugen. Somit besaßen die größten Vertreter der Moas, die in älteren Rekonstruktionen mit aufrechtem Hals gezeigt wurden und somit höher als ein Afrikanischer Strauß gewesen wären, Kopfhöhen von kaum mehr als zwei Metern. ⓘ
Die größten bekannten Exemplare von Eiern der Moas waren 40 Zentimeter hoch und hatten ein Gewicht von etwa 4500 Gramm, damit entsprach ihr Inhalt dem von mehr als 80 durchschnittlichen Hühnereiern. ⓘ
Fossilbericht
Der älteste Fund eines Moas im Fossilbericht ist Anomalopteryx aus dem späten Pliozän vor etwa 2,5 Millionen Jahren. Aus dem Pleistozän sind 33 fossile Überreste von Moas bekannt. Funde aus Ablagerungen vor dem Holozän sind demnach sehr selten, dies trifft jedoch für die fossile Überlieferung auf den neuseeländischen Inseln insgesamt zu. Alle bislang gefundenen Moa-Fossilien lassen sich den aus dem Holozän bekannten Arten zuordnen. Demnach sind während des Pleistozäns keine Moa-Arten ausgestorben oder entstanden, sondern lebten nahezu unverändert fort, bis sie beinahe gleichzeitig vom Menschen ausgerottet wurden. Es lässt sich lediglich oft eine leichte Größenabnahme zwischen dem Pleistozän und dem Holozän feststellen. ⓘ
Auch wenn entsprechende fossile Belege fehlen, sind die Moas eine weit ältere Tiergruppe, als ihr Fossilbericht es bislang dokumentiert. Von den Vorfahren der Moas konnten bislang keine Fossilien gefunden werden. ⓘ